

Champignons exotiques (suite)* le genre Hypsizygus ブナシメジ Buna-shimédji
La piste du « faux vrai Shimeji » conduit à une rarissime espèce de Bulliard !
par Guy FOURRÉ - 152, rue Jean-Jaurès - F - 79000 Niort (France) et Daniel GUEZ - Kita-Sumoto-cho 24-5 (Japon)
[4]
 Résumé
Résumé
Un champignon cultivé et présenté par les champignonnistes japonais sous le nom de « Honshiméji = Lyophyllum aggregatum » avait intrigué les auteurs : après enquête ils expliquent qu'il s'agit d'un faux Shiméji (alors que le préfixe hon veut dire « vrai » !) et qu'il n'a rien à voir avec Lyophyllum decastes (= L. aggregatum).
Le véritable Shiméji a été recombiné récemment par les mycologues japonais en Lyophyllum shimeji (Kawam.) Hongo, alors que l'espèce largement cultivée sous le nom de Honshimeji devrait être appelée Buna-shiméji et correspond à Hypsizygus tessulatus (Bull. : Fr.) Singer (= H. marmoreus (Peck) Bigelow.). C'est une espèce voisine de Hypsizygus ulmarius (Bull.: Fr.) Redhead (= Lyophyllum ulmarium (Bull.: Fr.) Kühn.), bien connue en Europe. L'auteur américain P. Stamets attribue aux Hypsizygus (H. tessulatus mais aussi H. ulmarius) des vertus insoupçonnées, tant sur le plan culinaire que thérapeutique. Décrit à l'origine par Bulliard, H. tessulatus n'était connu qu'en Amérique du Nord, mais il a été retrouvé récemment dans la nature, en Italie.
Dans le numéro d'avril 1993 de ce même bulletin, l'un de nous (GF) avait publié un article d'une douzaine de pages sur les « Champignons exotiques », article basé essentiellement sur les cultures insolites réalisées par un biologiste chinois travaillant en France, le Dr Guanglaï Zhang, des Ets. Le Champion à Beaulieu-les-Loches (Indre-et-Loire).
Cet article fut le point de départ de très fructueux échanges avec DG qui n'est pas un inconnu pour nos lecteurs', [...] a l'avantage d'être à la fois mycologue et japonisant. Après avoir pris connaissance du numéro d'avril 1993, il avait écrit à GF pour donner des compléments d'information et répondre à certaines questions posées dans l'article publié. Dès cette époque, nous avions formé le projet d'écrire en commun une suite nipponne (ni mauvaise) à ces « champignons exotiques » [...]. Mais, entre temps, le voyage de l'un de nous (GF) dans l'Ouest des U.S.A. nous a permis d'obtenir des informations complémentaires sur des champignons fort intéressants, dont une espèce qui existe chez nous mais semble méconnue.
-------------------------------------------
* Cet article est une suite de celui qui avait été publié, sous le même titre général « Champignons exotiques », dans notre numéro 129 (avril 1993).
1 Il avait publié dans le numéro de janvier 1990 (116:12-14) de notre bulletin un article de plusieurs pages, sous le titre Aperçu sur la flore mycologique du Japon.
[5]
L'un des principaux problèmes à résoudre était l'identité réelle du « Honshimeji », cultivé en France par M. Zhang, et vendu depuis longtemps par les champignonnistes japonais. Lors de la visite de l'un de nous (GF) au Stand du Japon au Salon International de l'Alimentation à Paris, en octobre 1988, le technicien japonais qui nous avait remis une documentation (en anglais et en français) sur le « Honshimeji » nous avait dit que le nom scientifique était Lyophyllum aggregatum, ce que nous avions traduit, pour respecter les noms valides, par Lyophyllum decastes. M. Zhang le présentait également sous ce binôme.
Dans notre article de 1993, nous avions écrit dès les premières lignes : « Mais... est-ce bien le même que chez nous ? ». Les fructifications que nous avions observées et photographiées, au laboratoire de M. Zhang puis à notre domicile, sur du substrat ensemencé par le spécialiste chinois, semblaient en effet sensiblement différentes de notre Tricholome agrégé, notamment par la présence très frappante d'une ornementation marbrée sur la cuticule, à tous les stades d'évolution. Les caractéristiques organoleptiques nous laissaient également songeur, le Honshimeji semblant considéré comme l'un des plus savoureux champignons, ce qui n'est point l'appréciation la plus couramment portée sur notre Tricholome agrégé. Enfin le nom « Honshimeji » semblait avoir été employé, par les Japonais eux-mêmes, pour désigner au moins deux espèces différentes, car un article du Japan Times d'octobre 1992 annonçait que l'on espérait réussir bientôt à cultiver ce champignon, alors que 4 ans plus tôt, au SIAL de Paris, il était présenté comme une production habituelle et parfaitement maîtrisée !
Les relations de DG avec les mycologues nippons et l'accès aux textes japonais originaux allaient permettre d'éclaircir ce mystère. D'abord M. Zhang s'était trompé en traduisant « Honshimeji » par «faux mousseron », c'était exactement le contraire : « hon » signifie « vrai ». Et « shimeji » désignait au Japon, dans le langage populaire, un champignon à lamelles, délectable, mais très imprécis. Il est mentionné dans des textes du 17e siècle et des estampes le figurent avec un port effectivement tricholomoïde. Mais en-dehors des mycologues, de quelques mycophages privilégiés et de certains gastronomes fortunés, la plupart des Japonais ne le connaissent que par ouï-dire, parce que les grand-mères en parlaient avec éloge. Tout Japonais connaît le vieil adage : « Matsutaké odeur, Shiméji saveur ».
Les champignonnistes ont achevé d'embrouiller les choses en nommant « HonShimeji » (« vrai Shimeji » !) une espèce voisine des Lyophyllum, qui a envahi les supermarchés nippons à partir de 1984 : c'est celui dont il était question dans les documentations distribuées au SIAL en 19882, et également celui que nous avions et photographié au laboratoire de M. Zhang. Mais il s'agit d'un «faux vrai Shiméji » ! (nous avons bien eu en France de « faux vrais passeports » !). Il est probable, d'ailleurs, que M. Zhang avait voulu exprimer cette idée de « faux shimeji », car il ne pouvait ignorer que « hon» signifie vrai.
Pour mieux faire comprendre cet embrouillamini, on pourrait comparer le terme « Shiméji » à notre « mousseron », qui n'est guère plus précis, mais qui évoque également, dans nos campagnes, le souvenir d'un champignon délicieux. Imaginez que nos champignonnistes,
---------------------------------------------------
2. Il s'agissait d'une usurpation du nom japonais d'une espèce renommée, pour en faire vendre une autre, cultivée mais inconnue du public. Cette substitution suscita d'ailleurs les protestations des mycologues japonais. De plus, à l'exportation, les champignonnistes japonais utilisaient pour la documentation commerciale le nom scientifique le plus familier aux occidentaux (aggregatum), alors que cela ne correspondait plus à la taxonomie japonaise moderne.
lorsqu'ils ont commencé à cultiver le Pleurotus ostreatus, l'aient appelé « vrai mousseron » pour en faciliter la commercialisation, et nous serions dans la même situation que les Japonais avec leur « faux vrai Shiméji »! L'auteur américain Paul Stamets, dont nous reparlerons plus loin, écrit lui aussi que « le nom « Shimeji » est utilisé dans un sens très large pour désigner quelques uns des meilleurs champignons japonais, et qu'il a été attribué à une vingtaine d'espèces différentes, suscitant une grande confusion chez les profanes...»
Quant aux mycologues japonais, ils ont d'abord considéré, dans le passé, le Shiméji comme un Tricholome endémique (Tricholoma shimeji Kawamura), puis il fut assimilé par Hongo à Tricholoma aggregatum (Schaeff.) Costantin & Dufour, puis à Lyophyllum aggregatum (Secr.) Kühner = L. decastes (on retombe ici sur le nom qui nous avait été donné au Sial à Paris pour une autre espèce), mais ils semblent maintenant s'être mis d'accord sur l'identité du « vrai vrai Shiméji », en adoptant un nouveau taxon: Lyophyllum shimeji (Kawam.) Hongo. C'est celui dont les tentatives de culture avaient fait l'objet de l'article du Japan Times en octobre 1992, et qui serait mycorrhizien (d'où la difficulté de le cultiver).
Après l'information publiée en 1992 dans le Japan Times sur les tentatives de domestication du « Hon-shimeji », un nouvel article paru dans Asahi Shinbun du 19/9/93, signalait que « l'Institut de recherches Yamato-Mycologie (département de Nara), qui commercialise également les blancs de culture, déclare avoir réussi pour la première fois la culture du Hon-Shimeji ». Et le journal japonais mettait les choses au point en ajoutant : « Les champignons de culture vendus dans les supermarchés sous le nom de Shimeji ne sont en réalité que des Hirataké (Pleurotus ostreatus) ou des Buna-shimeji (Hypsizygus marmoreus)3. Ce vrai Shimeji devrait être commercialisé en février prochain et vendu à un prix inférieur à celui du Shimeji naturel qui avoisine 600 F/kg ».
Nous ne nous attarderons pas d'avantage sur ce vrai-vrai Shimeji, que nos lecteurs n'ont guère de chances de rencontrer dans la nature, et qui n'est sans doute pas près d'apparaître sur nos marchés, même si les chercheurs japonais en maîtrisent maintenant la culture. Il est proposé, en février 1996, dans un supermarché de la banlieue d'Osaka, au prix de 400 F/kg.
Un Hypsizygus voisin de ulmarius...
Mais il restait à identifier le champignon que nous avions vu et photographié en France, cultivé et vendu sous le nom, usurpé, de Hon-Shiméji, et pour lequel les mycologues japonais avaient proposé le nom de Buna-Shiméji (Buna = hêtre), sans être suivis par les champignonnistes.
Cette espèce avait d'abord été englobée, par les mycologues japonais, dans un Lyophyllum ulmarium sensu lato, avant que Tsuguo Hongo et Heinz Clémençon ne la distinguent sous le nom de Lyophyllum ulmarium (Bull. : Fr.) Kühner, sensu auct. jap. p.p.
------------------------------------------------------------------------------------------------
3. DG signale que, depuis peu, il a pu constater dans les supermarchés japonais un étiquetage plus scrupuleux que dans le passé, avec le nom auquel les consommateurs sont habitués — même s'il est usurpé — mais suivi du nom scientifique japonais: «HONSHIMEJI (Buna-shiméji) pour l'Hypsizygus marmoreus = tesselatus cultivé depuis longtemps; SHIMEJI (Hira-také) pour Pleurotus ostreatus de culture; et AJI-SHIMEJI (Hon-shiméji) pour Lyophyllum shimeji (le vrai Shiméji). Ce dernier nom commercial est génial (et honnête), car ce sont les deux premiers mots de l'adage «aji shimeji, matsutake kaori» (shimeji pour la saveur, matsutaké pour l'odeur), dont l'impact médiatique est très important au Japon.
[7]
Il restait encore à lui trouver un nom lorsque Singer vint mettre fin à une nouvelle confusion en proposant, pour le « Buna-shimeji », le taxon Hypsizygus marmoreus (Peck) Bigelow (= Pleurotus elongatipes Peck).
Voici la traduction de la description de Tsuguo Hongo, dans le Hoikusha4, pour Hypsizygus marmoreus (Peck) Bigelow :
- Chapeau : 4, 5 - 15 cm, hémisphérique ou un peu dissymétrique, blanchâtre à crème légèrement brunâtre, présentant souvent des marbrures au contour peu marqué, la marge étant plus pâle.
- Lames : adnées-arrondies au pied, blanchâtres, peu espacées à serrées.
- Chair : épaisse, blanche, odeur plus ou moins farineuse.
- Pied : 3-10 x 0,6 - 2 cm, central ou excentrique et souvent courbe quand il pousse à l'horizontale, égal ou quelque peu fusiforme, concolore ou presque blanc, pubescent à la base.
- Spores : largement ovoïdes à subglobuleuses, 4-5 x 3,5-4 µm.
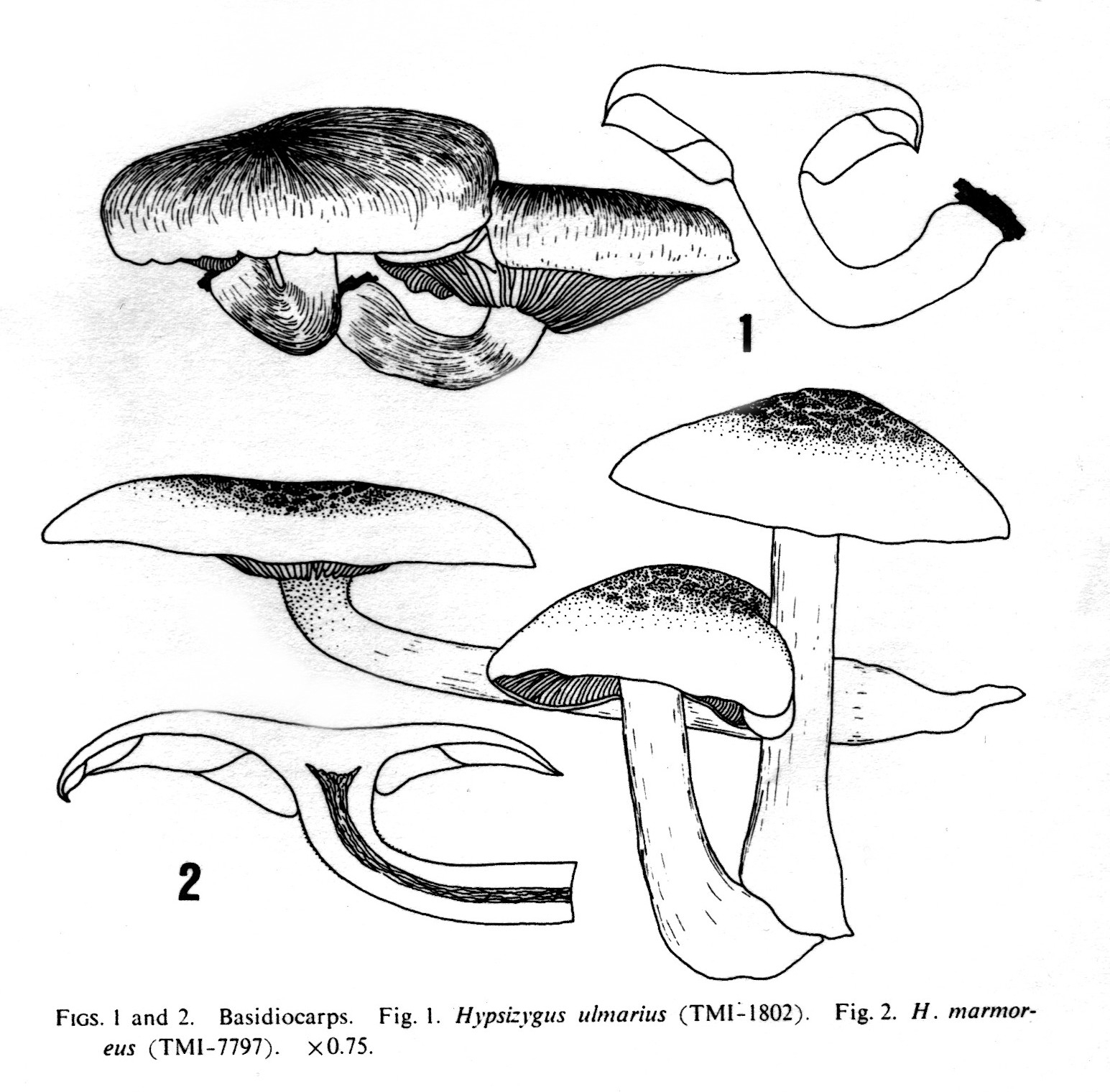
- Vient en troupes en automne sur les troncs morts ou vivants de divers feuillus (surtout Fagus et Acer). Zone tempérée de l'hémisphère nord. Bon comestible. Cultivé sous les noms de Hon-Shimeji ou Takara-Shimeji (Takara = trésor, N.D.T.). Se distingue de Hypsizygus ulmarius (Bull.:Fr.) Redhead par les marbrures qui ornent le plus souvent le chapeau, et par une spore inférieure à 5 pm. H. ulmarius possède un chapeau blanchâtre à brunâtre, plus ou moins craquelé, et une spore supérieure à 5 pm.
Taxonomie:
Le genre Hypsizygus a été créé par Singer en 1947 pour y verser les Tricholomataceae s'éloignant :
1) des Lyophyllum par l'absence de granulations sidérophiles dans les basides et de pigment incrustant ;
2) des Clitocybe par les lames non décurrentes, l'habitat lignicole et la forme des spores ;
3) des Tricholoma par l'habitat lignicole et le stipe non central ;
4) des Pleurotus par les lames non décurrentes, l'absence de cheilocystides et la forme des spores.
Le genre Hypsizygus, ainsi limité, ne contiendrait à ce jour que deux espèces : Hypsizygus ulmarius (Bull.:Fr.) Redhead (= Lyophyllum ulmarium (Bull.:Fr.) Kühn.), considéré comme type du genre ; et Hypsizygus tessulatus (Bull.:Fr.) Sing. (= H. marmoreus (Peck) Bigelow), qui n'était connu dans le passé que d'Amérique du Nord.
A propos du genre Hypsizygus, Stamets note qu'il s'éloigne de Lyophyllum également par l'écologie: il s'agit d'espèces poussant plutôt sur les troncs d'arbre (Hypsizygus signifierait «assemblés en l'air, ou en hauteur»)
-----------------------------------------------------------
4 A propos des Atlas japonais, il est bon de préciser, pour nos lecteurs européens, qu'il existe là aussi une confusion entre deux ouvrages récents désignés tous les deux sous le nom de Atlas d'lmazeki et Hongo, alors que ce vénérable premier auteur japonais, né en 1903, n'a guère participé ni à l'un ni à l'autre. Le principal auteur réel est Tsuguo Hongo, et il s'est fait assister, pour les dernières éditions, par d'autres mycologues qui sont à peine cités. Mais les Japonais ont un tel respect des anciens qu'ils ne publieraient pas un livre sans mettre en premier le nom de leur doyen !
Cet atlas a été publié sous deux formes: un ouvrage en deux volumes, équivalent de la flore de Kühner et Romagnesi ou de Moser pour les mycologues japonais, édité par Hoikusha; et une « bombe photographique », qui est l'illustration de la flore, avec d'excellentes photos d'Izawa, et des diagnoses résumées. Cet atlas photographique a été édité par Yama-Kei, et c'est celui qui est le plus diffusé en Occident. Pour éviter toute confusion, les mycologues japonais désignent ces deux ouvrages par le nom de l'éditeur, c'est-à-dire « le Hoikusha » ou «le Yama-Kei », et ils utilisent surtout le premier.
[8] [9]

[10]
alors que les Lyophyllum sont plutôt terrestres. Les Hypsizygus seraient ainsi plus proches des Pleurotes par leur habitat.
En remontant à l'origine de cette stirpe passablement confuse, on trouve un Agaricus tessellatus décrit en 1791, d'Europe, par Bulliard, dans « Histoire des champignons». Le basionyme est accompagné d'une planche en couleurs (513, 1), avec la légende suivante: « Cet Agaric se trouve en automne sur de vieilles poutres de chêne et quelquefois aussi sur de vieux troncs de pommier. Il est très agréable au goût et à l'odorat, mais un peu coriace5 . »
Fries le transforme en Pleurotus tessulatus (Systema mycologicum, 1821). Persoon, dans Mycologia europaea (1825), cite les deux taxons Agaricus tessellatus Bull. et Pleurotus tessulatus Fr., avec les descriptions originales, sans autres annotations ni commentaires,.
En 1872, Peck décrit une forme plus pâle sous le nom d'Agaricus marmoreus. Ricken (1915) cite tessulatus comme une simple variété de ulmarius. Kühner et Romagnesi (1953) évoquent tessulatus dans une note à propos d' ulmarius. Après la création du genre Hypsizygus par Singer en 1947, Bigelow transfére dans le nouveau genre le taxon de Peck, qui devient ainsi Hypsizygus marmoreus (Peck) Bigelow. Moser (1978) donne deux espèces dans le genre Hypsizygus, tessulatus (Bull.) Sing. et circinans (Fr.) Sing., différant par l'ornementation du chapeau, la couleur et l'insertion des lames (légèrement décurrentes pour circinans). Watling et Gregory (1989) retiennent dans le genre Hypsizygus ulmarius et H. tessulatus, divergents par leurs dimensions, l'odeur et l'ornementation de la surface piléique. Sur ce dernier point, Redhead (1986), souligne que l'ornementation « tesselée » de l'espèce est innée, très nette même sur les exemplaires jeunes, alors que pour ulmarius au contraire, une cuticule entière au départ se dissocie parfois en aréoles avec la croissance. Redhead sépare donc les deux espèces d'après l'ornementation piléique et les dimensions différentes des spores.
Pour Stamets, Hypsizygus tessulatus est synonyme à la fois de Hypsizygus marmoreus et de Pleurotus elongatipes, que les Américains appellent « the Long Footed Oyster Mushroom ( le Pleurote en huître à long pied) (Redhead 1986). Stamets ajoute que « en raison de la confusion associée au nom d'espèce H. tessulatus, les mycologues japonais préfèrent utiliser le nom taxonomiquement plus clair de H. marmoreus (Peck) Bigelow ».
Ainsi Nagasawa et Arita (1988) confirment, d'après des récoltes effectuées au Japon pour les deux espèces, la séparation des Hypsizygus entre H. ulmarius (Bull. : Fr.) Redhead, qui est le «shiro-tamogui-také « des Japonais; et H. marmoreus (Peck) Bigelow (= Pleurotus elongatipes Peck, = H. tessulatus (Bull. : Fr.) Sing ss Redhead, = Agaricus tessellatus Bull.), ou encore « Bunashimeji » pour les mycologues nippons et.... faux « vrai Shiméji » pour les champignonnistes! Ouf!
----------------------------------------------------------------------------------------
5 Nous remercions notre ami Jean Mornand, qui a assuré pour nous la reproduction de la planche de Bulliard.
E. Ponzi (1993), attire l'attention sur la transformation, probablement par Fries, du tessellatus de Bulliard en tessullatus, cette dernière orthographe ayant été reprise depuis par tous les auteurs. Ponzi rapporte l'avis d'un latiniste, pour lequel les deux termes pouvaient avoir la même signification. De sorte que la variation lexicale ne serait pas vraiment une erreur. Pour respecter le Code international de nomenclature, Singer aurait dû revenir à l'orthographe du basionyme et écrire tessellatus et non tessullatus. Redhead (1986) écrivait d'ailleurs correctement tessellatus.
[11]
Les vertus insoupçonnées des Hypsizygus
En août 1994, au congrès mondial de mycologie à Vancouver, l'un de nous (GF) avait rencontré le spécialiste américain Paul Stamets, qui produit et vend du mycélium d'espèces tout à fait surprenantes (pour nous). Après le congrès, nous étions passé lui rendre visite à son laboratoire, à Kalmiche, près d'Olympia, dans l'Etat de Washington, tout près de l'Océan Pacifique (et non loin de Seattle). Il nous avait alors remis son dernier ouvrage, Growing gourmet and medicinal Mushrooms (1993), qui compte plus de 500 pages, dont 13 consacrées aux Hypsizygus, ces champignons possèdant à ses yeux des vertus tout à fait remarquables.
Paul Stamets décrit donc la culture des Hypsizygus (tessulatus et ulmarius) et il estime que ces champignons, par leur chair plus ferme, surclassent, sur le plan gustatif, toutes les espèces ou variétés de Pleurotes cultivées en Europe et en Amérique du Nord. Pour les champignonnistes, un autre avantage par rapport aux Pleurotes est que les Hypsizygus produisent beaucoup moins de spores8. De plus, le mycélium de H. tessulatus élabore des substances toxiques pour les nématodes, et Paul Stamets confirme qu'il n'a jamais constaté la présence, dans ses cultures de «Buna-shimeji», de ces petits vers indésirables.
 Récolte de Buna-shimeji en foret de Kusabi-Iwa (Miyoshi, Hiroshima) en octobre 2020 DG
Récolte de Buna-shimeji en foret de Kusabi-Iwa (Miyoshi, Hiroshima) en octobre 2020 DG

Hypsizygus tessulatus (Bulliard : Fries) Singer (= H. marmoreus (Peck) Bigelow, est une espèce connue dans la nature en Amérique du Nord (ainsi qu'en Europe et en Asie selon Stamets), où elle existe sur orme, hêtre, peuplier, érable, saule, chêne, etc. Elle peut avoir un comportement parasite sur les troncs malades, ou saprophyte sur les arbres morts. Des souches utilisables sont facilement obtenues à partir des fructifications récoltées dans la nature. La culture est facile, par inoculation de bûches ou de souches, sur des couches de copeaux de bois ou de sciure. Paul Stamets indique même dans son ouvrage les techniques à employer pour obtenir le mycélium. Il démontre que les grandes différences de longueur du pied, qui ont intrigué dans le passé des mycologues américains visitant des exploitations au Japon, résultent des variations des taux de lumière et d'oxyde de carbone. En réduisant la lumière et en augmentant le taux de CO2, on provoque l'élongation des stipes pour créer des bouquets émergeant de bouteilles à goulot étroit !
Sur la valeur culinaire de ce champignon, l'auteur américain ne tarit pas d'éloges : «c'est une espèce comestible supérieure, avec une texture ferme, croustillante, et une douce saveur de noisette; ce champignon est hautement estimé au Japon. Polyvalent et mettant en valeur n'importe quel plat, H. tessulatus s'accorde très bien avec des légumes, de la viande, des fruits de mer ou fritures, et il peut être ajouté à des soupes ou sauces à la dernière minute de la préparation. L'étendue de la saveur de ce champignon est difficile à décrire. Buna-shimeji peut être incorporé dans un large choix de recettes».
Ce n'est pas tout : ses vertus médicinales pourraient être encore plus intéressantes. Selon Paul Stamets, « Une étude non publiée provenant de l'Institut
---------------------------------------------------------------------------------------------
C'est aussi l'opinion de DG, qui place le Buna-shimeji au-dessus du shii-také, pourtant excellent. Au Japon, il remplace de plus en plus le champignon de Paris dans les plats « étrangers » les plus divers, alors que le shii-také conserve sa place dans la cuisine traditionnelle nipponne. Sa culture maîtrisée le rend de plus très bon marché.On sait que l'énorme production de spores de pleurotes provoque parfois des allergies dont sont victimes les employés des champignonnières, à tel point que L'INRA de Bordeaux a tenté de mettre au point des souches de Pleurotus ostreatus ne produisant pas de spores.
[12]
National du Cancer au Japon, par Ikekawa (1990), montre une forte activité anti-tumorale. Des souris avaient été inoculées avec le Carcinome de Lewis Lung, et on leur donnait des extraits aqueux du champignon frais. A une dose équivalente à 1 g/kg de poids corporel, par jour, les tumeurs étaient inhibées à 100%, et il s'ensuivait une totale régression. Des groupes témoins confirmaient que, sans l'extrait de champignon, les tumeurs continuaient à croître. Aucune étude sur les humains n'a encore été signalée».
Il ne faut pas rêver, on n'en est pas encore, en Occident, à soigner les cancers avec des extraits de ces champignons. Mais le livre de Paul Stamets contient de très nombreuses références de ce genre, pour une grande variété d'espèces que l'on pourrait récolter ou cultiver chez nous, et dont on fait grand cas en Asie.
Pour en revenir à la valeur gustative, Paul Stamets attribue de même une excellente note à Hypsizygus ulmarius (qu'il cultive également), et qu'il trouve bien supérieur aux Pleurotes. Les seules espèces qui pourraient lui être favorablement comparées seraient justement son «cousin» H. tessulatus et, dans la nature, Pleurotus eryngii... (cette dernière comparaison pourrait constituer une référence pour les mycophages européens). Avec un avantage supplémentaire, celui de chapeaux pouvant atteindre une grande taille.
Mais la valeur gustative de notre ancien Lyophyllum uimarium (devenu Hypsizygus) semble bien méconnue en Europe : il n'est même pas cité dans la liste des cent onze espèces de qualité énumérées par Paul Ramain, ni dans les 135 de Roger Heim, ni dans les 114 de Umberto Nonis9. Il est vrai qu'il s'agit d'une espèce peu commune, mais on peut se demander aussi s'il n'existe pas des différences sensibles, sur le plan des caractéristiques organoleptiques, entre des souches de champignons botaniquement semblables, mais fructifiant dans des pays très éloignés les uns des autres nous nous sommes déjà posé la question à propos du Tricholoma caligatum de notre pays, théoriquement identique au Matsutaké des Japonais.
Le Buna-shiméji récolté _dans la nature en Italie!
Si Hypsizygus ulmarius est bien connu des mycologues de notre pays, il n'en est pas de même de l'autre espèce Hypsizygus tessulatus, dont il n'a guère été question en Europe depuis Bulliard...
Mais en septembre 1993, la revue italienne Rivista di Micologia publiait un article de Enrico Ponzi, de Parma, signalant plusieurs récoltes de Hypsizygus tessulatus (Bull.: Fr.) Sing. - le Buna-shiméji cultivé par les Japonais ! - dans la nature. L'auteur rapporte qu'il a vu pour la première fois cette espèce à l'exposition du groupe G. Passerini de Parme, en octobre 1990, la récolte ayant été effectuée à Ozzano (PR, Italie) sur souche de chêne par R. Roberti et P.L. Pernis. L'espèce fut retrouvée en octobre 1991 à Gotra (PR) par E. Ponzi et G. Guasti, à nouveau sur souche de chêne.
------------------------------------------------------------------------------------------------------
9 Cette liste est reproduite dans l'Atlas de Rinaldi & Tyndalo, Ed. Nathan, 1973.
10 DG confirme que la valeur organoleptique du Matsutaké est très variable selon son origine: les importations de Corée sont loin d'avoir le parfum suave de leurs homologues nippons. Il est vrai que l'hôte est différent (Pinus koraiensis). Au Japon même, les « variétés » qui ne poussent pas sous Pinus densiflora (Aka-matsu) sont sans valeur commerciale et sont affublées de noms péjoratifs. Il ne fait pas de doute qu'il s'agit de « souches organoleptiques » distinctes, l'odeur étant ici le critère distinctif.
[13]
Par rapport à H. ulmarius, les mycologues italiens ont remarqué des dimensions piléiques semblant inférieures, une forte odeur de farine fraîche, perceptible même quand le champignon se trouve encore sur le substrat, et une ornementation tesselée, plus marquée sur les jeunes exemplaires, mais persistante même sur les carpophores complètement développés.
Les seules divergences observées, par rapport aux descriptions de tessulatus, auraient été la plus grande longueur du stipe et l'habitat sur souche morte alors qu'il était décrit comme parasite des arbres vivants. Mais l'auteur italien estime, à juste titre, que ces divergences peuvent facilement s'expliquer, la longueur du stipe par une croissance dans une cavité obligeant le champignon à s'allonger pour atteindre un espace dégagé, et l'habitat sur souche morte par le fait, bien connu, que beaucoup d'espèces parasites comportent dans leur cycle biologique une phase saprophyte. Le comportement alternativement parasite ou saprophyte de Hypsizygus tessulatus est d'ailleurs également mis en évidence par l'Américain Stamets.
Enrico Ponzi développe dans son article les problèmes de nomenclature (nous y avons trouvé une partie des éléments rapportés ci-dessus au chapitre de la taxonomie"), puis il explique les différences de couleur de la pourriture provoquée par le mycélium de cette espèce, selon les structures attaquées, cellulose ou lignine.
Il signale enfin que, sauf erreur, cette espèce n'avait pas encore été trouvée en Italie, et qu'il n'en existe guère d'illustration valable dans les ouvrages des auteurs modernes: une planche de Dermek (1987) portant le nom de Hypsizygus tessulatus représente en fait très clairement... H. ulmarius.
Enrico Ponzi publie une photo en couleurs (par Guasti) de la récolte de H. tessulatus faite en Italie. On y retrouve de façon très frappante la culticule marbrée qui avait retenu notre attention dans les cultures de notre Chinois de France.
Ainsi la boucle est bouclée : depuis Paris (SIAL de 1988) et les Ets Le Champion à Loches en Touraine (1992), en passant par le Japon et les USA (1994), la piste du «faux vrai Shiméji» nous a conduit à cet Agaricus tessellatus de Bulliard, champignon bien oublié dans notre pays. Mais la récolte effectuée en Italie semble démontrer que cette espèce rarissime, connue seulement d'Amérique du Nord depuis des lustres, existe aussi en Europe. Bulliard n'avait pas rêvé!
ADDENDA — CORRIGENDA À L'ARTICLE DE 1993
Par sa connaissance de la langue japonaise, et ses relations avec les mycologues nippons, l'un de nous (DG), a pu corriger ou compléter certaines informations figurant dans l'article que l'autre (GF) avait publié en 1993 dans ce même bulletin. De plus, d'autres informations ont été obtenues depuis, notamment par les publications du spécialiste américain Paul Stamets. Voici ces compléments ou corrections
Pleurotus salmoneostramineus L. Vassiljeva
La description de cette espèce dans ce que nous appelions «l'atlas d'Imazeki »4 a été rédigée en fait par Tsuguo Hongo, le nom d'Imazeki n'apparaissant en premier que pour des raisons «honoris causa». Nous avions publié une traduction en français de la description de Pleurotus salmoneostramineus, mais cette traduction comportait une erreur : c'est le pied et non le chapeau qui est orné d'un fin duvet blanchâtre, la cuticule étant glabre.
---------------------------------------------------------------------------------------------
11 Nous remercions vivement le Dr Vincenzo Miggliozzi (Roma), qui nous a aidé à traduire de l'italien les paragraphes concernant la nomenclature.
[14]
 L'article du Japan Times en date du 30/9/90 annonçant le lancement de la culture du Pleurote rose concernait en fait la commercialisation sur une grande échelle de cette espèce, qui était cultivée depuis plus de 10 ans par Nagasawa et son équipe au «Tottori Mycological Institute ».
L'article du Japan Times en date du 30/9/90 annonçant le lancement de la culture du Pleurote rose concernait en fait la commercialisation sur une grande échelle de cette espèce, qui était cultivée depuis plus de 10 ans par Nagasawa et son équipe au «Tottori Mycological Institute ».
Nous avions mis en doute en 1993 la synonymie proposée par Delmas (1989) entre Pleurotus eous (M.J. Berk.) Sacc. et Pleurotus salmoneostramineus L. Vassiljeva. L'Américain Paul Stamets mettrait tout le monde d'accord en considérant que ces deux taxons, de même que P. flabellatus (Berk. & Br.) Saccardo, P ostreato-roseus Singer et P djamor var. roseus Corner, font partie intégrante d'un complexe de formes sub-tropicales qui pourraient être nommées Pleurotus djamor (Fries) Boedjin sensu lato, le basionyme étant le Lentinus djamor de... Fries (1838).
Sur les variations de couleurs des lames et de la sporée, Paul Stamets affirme que le rose plus ou moins intense des lames dépend de l'âge mais aussi des souches, et que la couleur des spores dépend de celles des lames (ou vice-versa !). On peut ainsi obtenir des lames et spores roses ou blanches, parfois à partir des mêmes dicaryons! L'auteur américain estime que les Pleurotes roses représentent un vaste complexe de variétés atteignant un stade très rapide d'évolution convergente ou divergente, et que des recherches de DNA seront sans doute nécessaires pour éclaircir la taxonomie de ce groupe.
Paul Stamets souligne enfin que c'est de loin l'espèce la plus agressive, la plus apte par conséquent à s'adapter à des conditions précaires de culture, et à coloniser un substrat non pasteurisé à une vitesse record, les fructifications pouvant apparaître dix jours seulement après la mise en place du mycélium. Des avantages que nous avons pu vérifier à maintes reprises, et qui sont particulièrement importants pour les amateurs ne disposant que de moyens de culture rudimentaires. Enfin nous sommes d'accord avec l'appréciation de Stamets sur la valeur culinaire de ce Pleurote rose, qui est le plus beau, mais pas le meilleur sur le plan gastronomique, la chair étant un peu plus coriace que celle des autres Pleurotes de culture.
Auricularia polytricha (Mont.) Sacc.
«Kikurage» (phonétiquement Ki-Koura-Gué) désigne au Japon le genre Auricularia et plus particulièrement l'espèce auricula-judae. « L'oreille de Judas chinoise » (Auricularia polytricha) est appelée « Aragué-Kikuragué », ce qui signifie à peu près « méduse des arbres hirsute ».
Flammulina velutipes (Curt. :Fr.) P. Karst.
Le nom japonais Enoki (E-no-ki) est celui d'un arbre, plus précisément un micocoulier, Celtis sinensis var. japonica et « také » est le terme général pour les champignons. La « petite flamme » n'est donc pas la traduction du nom japonais, mais tout simplement celle du nom latin du genre.
Agrocybe chaxingu Huang
L'aire de répartition de cette espèce est probablement très limitée, car elle semble tout à fait inconnue des mycologues célèbres sur le plan mondial, y compris des Japonais ! Pour préciser sa région d'origine, nous avions écrit en 1993 une fois «Fujin» et une fois «Fujian»: ce n'est pas la même chose, et c'est la seconde qui est la bonne.
Voici la diagnose latine de Agrocybe chaxingu Huang (1973):
Fructificatio solitaria vel caespitosa, pileo carnoso 4-18 cm (generatim 8 cm)
[15]
diametro, primum convexo-campanulato, demum applanato, centro leviter depresso, non viscido pallide cinnamomeo, margine picto, dilutiore incurvatoque, post demum applanato, caro albo 1-2 cm crasso, lamellis adnatis, albis usque brunneis, 72-80 pm latis. Stipitibus a basi usque ad apicem subaequicrassis, 4-10 cm longis, 0,5-2 cm crassis, annulis membranaceis, basidiosporis ellipsoideis, 611,2 x 4,5 1.1M.
Fujian: Sanming. In trunco Camelliae oleiferae. VII 1970. Huang Nian-lai. N. 1 (Typus in San Ming Institutum Mycologicum).
Le texte du Dr Zhang, que nous avions publié en 1993, n'est pas la traduction de la diagnose originale en chinois, mais une description personnelle ou la traduction d'un autre texte.
Potentiel thérapeutique des champignons
Nous avions évoqué brièvement, dans notre article de 1993, la culture de certaines espèces à des fins exclusivement thérapeutiques, lorsqu'elles ne sont pas en même temps comestibles : ainsi le Ganoderma lucidum, dur comme du liège, n'a pas d'usage culinaire, mais en le faisant bouillir on en tire des tisanes bienfaisantes, ou on extrait les principes actifs de son mycélium en cultures submergées. Alors que le Shii-také (Lentinula edodes) est à la fois un excellent comestible (le meilleur de tous les champignons cultivés à notre goût) et un cocktail de « médicaments ».
Au congrès mondial de mycologie à Vancouver, les ressources thérapeutiques éventuelles offertes par les macromycètes ont fait l'objet de nombreux exposés, notamment pour Ganoderma lucidum, et il semble bien que les Américains aient emboîté le pas aux Asiatiques pour développer la recherche sur ces «champignons-médicaments». On peut regretter que l'industrie pharmaceutique, dans nos pays occidentaux, semble afficher un dédain souverain à l'égard de ces remèdes naturels.
Bibliographie
BULLIARD P., 1791 - Histoire des champignons de la France. Paris.
DELMAS J., 1989 - Les champignons et leur culture - Flammarion édit., Paris 970 p.
FOURRÉ G., 1993 - Champignons exotiques - Bull. Féd. Myc. Dauphiné-Savoie 129 4-17. FRIES E. M., 1821 - Systema Mycologicum. Lundae.
HEIM R., 1957 - Champignons d'Europe - Ed. N. Boubée & Cie.
IMAZEKI R. & HONGO T., 1987 - Colored illustrations of Mushrooms of Japan (en japonais). - Nouvel Atlas en couleur des champignons du Japon. Vol. 1 - Hoikusha Publ., Osaka : 316 p, 515 pl. (993 planches pour les 2 vol.).
IMAZEKI R., OTANI Y., HONGO T., 1988 - Fungi of Japan - Yama-Kei Publ., Tokyo - 624 p.
KÜHNER R. ET ROMAGNESI H., 1953 - Flore analytique des champignons supérieurs. Masson, Edit., Paris
MOSER M., 1978 - Die Rôhrlinge und Blâtterpilze - Stuttgart.
NAGASAWA E. & ARITA I., 1988 - Hypsizygus ulmarius - H. marmoreus - Rept. Tottori Mycol. Inst. 26 : 71-78.
PERSOON C., 1825 - Mycologia europaea. Erlangen.
PONZI E., 1993 - Hypsizygus tessulatus (Bull. : Fr.) Sing. - Un raro ritrovamento e un singolare equivoco nomenclatorio. Rivista di Micologia, XXXVI (3) : 245-250.
REDHEAD S.A., 1986 - Mycological observations 15-16 : on Omphalia and Pleurotus. Mycologia 78 : 522-528.
RAMAIN P., 1954 - Mycogastronomie. Les Bibliophiles Gastronomes, Paris.
RICKEN A., 1915 - Die Blâtterpilze. Lipsia.
SINGER R., 1947 - New genera of fungi. III - Mycologia 39: 77-89.
STAMETS P., 1993 - Growing gourmet and medicinal mushrooms. Fungi Perfecti, Edit., Olympia (USA) - 584 p., 452 ph.
WATLING R. & GREGORY N.M., 1989 - British Fungus flora 6 : Crepidotaceae, Pleurotaceae and other pleurotoid agarics - (Edinburgh).
1
Trans. mycol. Soc. Japan 25: 1-9, 1984
Mycological observations 13-14: on Hypsizygus and Tricholoma
S. A. REDHEAD
Biosystematics Research institute, Agriculture Canada, Ottawa, Ontario, Canada, K A OC6
(Accepted for publication 1 March 1984)
Key Word Index Hypsizygus; Tricholoma; Lyophyllum; Armillaria; American Matsutaké ;
nomenclature; biogeography; host plants.
Summary
The genus Hypsizygus Singer is accepted for two species, H. ulmarius (Bull.: Fr.) Redhead comb.nay. [syn. H. tessulatus (Bull.: Fr.) Singer] and H. marmoreus (Peck) Bigelow [syn. H. elongatipes (Peck) Bigelow]. Controversy concerning the identities of the North American and Eurasian collections of H. ulmarius is discussed with the conclusion that all are conspecific.
The name Agaricus magnivelaris Peck was noted to be the earliest validly published legitimate name for the American Matsutake. The combination Tricholoma magnivelare (Peck) Redhead comb. nov. is proposed to replace T. ponderosum (Sacc.) Singer. The species is documented from new localities in Canada.
This is the third contribution in a series of limited miscellaneous mycological observations (Redhead 1979, 1985). The distribution, taxonomy, and nomenclature of three common edible species are discussed.
13. Hypsizygus : The genus Hypsizygus Singer (1947) was proposed to accommodate Pleurotus ulmarius sensu North American authors in contrast to the European concept of Pleurotus ulmarius (Bull. : Fr.) Kummer which Singer recognized as Lyophyllum ulmarium (Bull.: Fr.) Kiihner. Singer (l.c.) indicated that the name Pleurotus tessulatus (Bull. : Fr.) Gillet applied to the North American species commonly labelled P. ulmarius and therefore designated P. tessulatus as type of the genus Hypsizygus. The name Pleurotus tessulatus is based on European materials seen by Bulliard (1791).
The supposed basis for separating the North American " P. ulmarius " from the European (?and Asian) " P. ulmarius " specifically and generically was the presence of carminophilous granules in basidia of the latter and their absence in the former (Singer 1947). Although not directly cited, Kühner's (1938) research pioneering the use of acetocarmine to stain basidia as a taxonomic guide laid the foundations for the recognition of the genus Hypsizygus. Kiihner (1938) noted that certain white-spored groups of Agaricales had basidia densely packed with darkly staining granules, whereas others lacked
[2]
these granules. The one exception was Pleurotus ulmarius. It was observed to have, "Des basides I distinctement granuleuses..." (Kuhner 1938 : 206). Although Singer and Kuthan (1980) recently tabulated suspected differences between P. ulmarius and P. tessulatus their arguments are not convincing. They noted a virtual absence of siderophilous [carminophilous] granules in the basidia, a distinct farinaceous odor, deeper cream coloured spore print, non-ventricose lower stipe, the absence of incrusting pigments in the pileipellis, and the preference for living or at least standing trees for the North American taxon. However, among the collections I have studied (cited below) there are collections from North America with ventricose lower stipes, with a distinct but not farinaceous odor, and varying low numbers (12-15) of distinctly carminophilous granules, particularly when stained while fresh. No differences were noted between the acetocarmine-stained basidia of the North American and European collections examined. The basidia are not densely filled with intensely staining granules as are most species of Lyophyllum and Calocybe and neither are they relatively unreactive as in other genera of the Tricholomataceae.
Much of the cytoplasm or at least large irregular globules are darkly but not intensely stained and a few smaller intensely stained granules can be seen. The latter appeared more abundant when fresh than after drying in at least one collection (DAOM 188211).
The European and Asian taxon regularly occurs on standing trunks of dead or living trees (Cooke, 1882; Lange, 1936; Dennis, 1981; Lundell and Nannfeldt, 1979; Imai,
1938 ; Pilat, 1935 ; Kawamura, 1954 ; Bresadola, 1927; Imazeki and Hongo, 1957) and the odor of the Eurasian taxon varies considerably from farinaceous to subfarinaceous, subnauseus or otherwise (1.c.). Eurasian and North American collections vary from being whitish to brownish on the pileus surface, and all degrees of cracking from mere fissures to areolate scaly, nearly squarrose scaly or even pyramidal scaly conditions were noted for North American collections. No differences were noted between spore print colours from North American collections and a print with Lundell and Nannfeldt (No. 2867).
Based on all of these observations I conclude that the North American Pleurotus ulmarius is conspecific with the Eurasian Pleurotus ulmarius and that the name Pleurotus tessulatus is based on a growth form of the species caused by environmental factors. Both basionyms, Agaricus ulmarius Bulliard and Agaricus tessulatus Bulliard were published simultaneously with coloured plates and with extremely abbreviated diagnoses (Bulliard, 1791). Both were treated as independent species by Persoon (1801) and Fries (1821). Quelet (1888) appears to have been the first to recognize them as specific synonyms, reducing A. tessulatus to varietal level. Thus the correct epithet is ulmarius (Art. 57.2, I.C.B.N.). Bulliard (1812) considered Agaricus palmatus Bulliard (1785) to be a synonym of A. ulmarius but this synonymy is not correct as A. palmatus is Rhodotus palmatus (Bull. : Fr.) Maire.
The generic position of Pleurotus ulmarius is debatable. In some aspects such as the Hypsizygus and TrichoIoma
[3]
mixed reaction of its basidia in acetocarmine stainit is intermediate between true Lyophyllums (with carminophilous spore walls) and staining basidial granules', and other Tricholomataceae or Polyporaceae lacking both [e.g. Pleurotus ostreatus ( Jacq.: Fr.) Pleurotus sensu stricto, Tricholoma, and Clitocybe, but not Lyophyllum. However, the tough consistency of the basidiomes and the ability to cause primary decay in livinK trees along with the unusually staining basidia separate P. ulmarius from other Lyophyllum species.
For these reasons I recognize an emended genus Hypsizygus as distinct from, but closely allied to Lyophyllum. The correct name for the type of this genus is Hypsizygus ulmarius (Bull.: Fr.) Redhead comb. nov. ; basionym Agaricus ulmarius Bulliard (1791: pl. 510).
Specimens examined (selected) :
Canada: Manitoba: Bruxelles, 7. VII. 1959, J. W. Groves (DAOM 65194) on Acer negundo L. ; Gilbert Plains, 20. VII. 1971, J. Parker 1149 (DAOM 188257) on living Populus balsamifera L., 7. IX. 1975, J. P. 5031 (DAOM 188258) on dead Populus 7 ft. off ground; Winnipeg, 6. XI. 1926, T. Johnson (DAOM 152331) ; also DAOM 157705, 152332, 155114. New Brunswick : Fredericton, 16. XI. 1954, W. R. Newell (DAOM 46371) on living Ulmus. Nova Scotia: Kentville, 9. XI. 1953, K. A. Harrison 2730 (DAOM 112190) on Ulmus. Ontario: Carleton place, 12. IX. 1891, J. Macoun (DAOM 51522) ; New Durham, 10. XI. 1930, R. F. Cain (DAOM 80607) on Ulmus; Ottawa, 13. X. 1923, 1. XI, 1922, W. S. Odell (DAOM 51521, 51519) on Ulmus, 2. X. 1930, I. Mounce (DAOM F1569) on Ulmus americanus L., 6. X. 1932, H. N. Racicot (DAOM F2418) on Acer negundo; 22. XI. 1983, S. A. Redhead (DAOM 188211) on living Ulmus glabra Huds. var. camperdownii Redh.; also DAOM F5767, 51525, 51526, F2115, F6993, 16545; Sand Point, 18. X. 1953, J. W. G. (DAOM 45167) ; Toronto, 30. X. 1918, W. S. 0. (DAOM 51520). Quebec: Cantley, Gatineau Co., 21. X. 1975, 23. X. 1978, J. H. Ginns 2977, 4133 (DAOM 154319, 170525) on Ulmus americanus; Poltimore, 11. X. 1981, J. E. & S. A. Redhead & A. & J. H. Ginns (DAOM 18108) on Acer negundo; Tenaga, 20. X. 1955, S. C. Thomson (DAOM 48725). Saskatchewan: Craven, 26. VII. 1960, M. K. Nobles (DAOM 71661) on Acer negundo; Indian Head, 27. VII. 1938, L. Petersen (DAOM 9174) on A. negundo; also DAOM 9173, F10184; Saskatoon, 11. XI. 1939, T. C. Vanterpool (DAOM 10888) on A. negundo; 6. X. 1951, C. G. Riley & P. J. Kristopovich (DAOM 28937) on Ulmus; also DA OM 40279. England: Edgware, 10. XII. 1959, ex K (DAOM 74812) ; Kew, ex K (DAOM F8072) on Ulmus. Sweden : Uppland, Upsala, 28. VII. 1946, A. Melderis (DAOM 65017) ; Vastergotland, Goteborg, 25. X. 1956 or 10. X. 1963, T. Karivall & A. Thungren (Lundell & Nannfeldt Fungi Exs. Suecici-2867, DAOM) on Ulmus. U. S. A.: Massachusetts:Belmont, 9. X. 1937, G. D. Darker 6508 (DA OM 96390) on Acer rubrum L.
4 S. A. REDHEAD
A closely related species, commonly labelled Pleurotus elongatipes Peck (1908), has often been confused with H. ulmarius. Many collections in DAOM originally labelled as P. ulmarius are in fact conspecific with P. elongatipes. Modern, descriptions of this species have been published by Smith (1957), Bigelow and Barr (1966) and Miller and Manning (1976). Miller (1972) published a coloured photograph. Bigelow and Barr (1966) noted that P. elongatipes probably belonged in the genus Hypsizygus. Following type studies, Bigelow (1970) later transferred Pleurotus elongatipes and Agaricus marmoreus Peck (1872)
to the genus Hypsizygus. Both differ from H. ulmarius by smaller spores which average under 5 pm in broadest dimension. Hypsizygus marmoreus (Peck) Bigelow, known from the type locality only, was differentiated from Hypsizygus elongatipes by the presence of decurrent lamellae. Peck (1872 : .61) reported the lamellae for A. marmoreus as, "close, narrow, arcuate, unequally decurrent..." For H. elongatipes Bigelow and Barr (1966 : 184) reported "Lamellae at first adnexed or adnate, becoming sinuate or emarginate...", Smith (1957: 56) reported "Lamellae adnexed becoming emarginate, rounded next to the stipe...", and Peck (1908: 1) reported, "lamellae thin, close, rounded behind, adnexed..."
On the basis of these observations two taxa could be recognized. However, when more collections are examined a broader range of variability in lamellar attachment becomes apparent, as does variability in other characters. A collection from Harwich, Kent Co., Ont. in the J. Dearness herbarium (DAOM) caused considerable consternation to both Dearness and C. C. Bell the collector. It was noted to resemble Pleurotus ulmarius or P. elongatipes but had decurrent lamellae, and also the smaller spores like P. elongatipes. Bell wrote to Dearness (litt. in herb.), "Gills:-adnate-decurrent to decurrent, somewhat close, whitish, edge entire, narrow (7 mm) ..." A sketch in the letter, and examination of the specimen revealed that the lamellae are unequally but not profoundly decurrent.
Another collection, by H. M. E. Schalkwyk 785 (DAOM 188232) exhibited a range of lamellar attachment from adnexed to adnate or adnate-decurrent. Other collections varied typically from profoundly adnexed to adnate. All are considered to be conspecific. One extremely abnormal basidiome (DAOM 160082) has profoundly decurrent lamellae, is extremely woody, and is broadly turbinate. The collector (H. M. E. S.) noted that it took 2 months to develop.
The more or less decurrent lamellar attachment noted above is an extreme in range of attachment not unlike that exhibited by Tricholomopsis decora (Fr.) Singer (cf. Smith, 1960). The oldest epithet for the taxon is Agaricus marmoreus, giving the combination Hypsizygus marmoreus (Peck) Bigelow priority over Hypsizygus elongatipes (Peck) Bigelow. In North America Hypsizygus marmoreus has a larger geographic range than H. ulmarius.
Whereas H. ulmarius is restricted to areas east of the continental divide and generally central latitudes, H. marmoreus is found coast to coast and north to the Yukon. Its affinity for Populus species in addition to Acer and Betula may explain its wider distribution. The species is not yet known from Europe. Pilat (1935) recognized the taxon only from North America under the invalid name " Pleurotus ulmarius form. burtianus ", published without Latin.
[5] Hypsizygus and Tricholoma
However, H. marmoreus probably occurs in Japan. An illustration of Lyophyllum ulmarium by Ito (1959, p. 92, fig. 38c [mislabelled as b] ) depicts a collection of a form like some H. marmoreus collections, and unlike H. ulmarius collections. The spores illustrated in fig. 38e also appear to be about 5 ,um diam. on average and more nearly globose than are most H. ulmarius spores.
Specimens examined (selected) :
Canada: Alberta: Calgary, 29. VIII. 1973, 21. VIII. 1978, R. M. Danielson 1134, 2808 (DAOM 150768, 184728) at base of Populus balsamifera; Sandy L., 72 km NW of Edmonton, 26. IX. 1976, 18. VII. 1977, H. M. E. Schalkwyk 723, 785 (DAOM 160995, 188232) on P. balsamifera. British Columbia : Glacier Natl. Park, Beaver R. Valley, 19. IX. 1980, S. A. Redhead 3095 (DAOM 187624) on large fallen Populus trichocarpa Torr. and Gray: Manning Prov. Park at Hope end, 5. X. 1962, M. Pantidou (DAOM 92244) ; Prince George, X. 1947, W. G. Ziller V-3098 (DAOM 21376) on butt of snag of P. trichocarpa; Quesnel, IX. 1947, G. P. Thomas V-2744 (DAOM 21272) on P. trichocarpa; Victoria, 14. X. 1963, M. C. Melburn 458 (DAOM 94387) on uprooted Abies; Winfield near Kelowna, X. 1980, ?leg. (DAOM 179146). Northwest Territories : Fort Smith, 17. VIII. 1950, Loan 264 (DAOM 25202) at base of Populus tremuloides Michx. Nova Scotia: Cold Brook, 14. X. 1932, K. A. Harrison 459 (DAOM 111009) on Populus; Kentville, 21. X. 1932, K. A. H. 460 (DAOM 111010). Ontario: Algonquin Prov. Park, Hardwood Hill trail, 3. IX. 1977, G. Thorn (DAOM 186652) on trunk of living Fagus grandifolia Ehrn. ; Byron bog near London, 17. X. 1925, J. Dearness (DAOM) on Ulmus; Dorset, 30. IX. 1950, C. G. McKeen (DAOM 26271) from 10 ft. canker on stand- ing Acer saccharum, 2. X. 1950, C. T. McK. (DAOM 25000) on cut log of A. saccharum; Fallowfield, 5. X. 1954, J. W. G. (DAOM 48056) ; Harwich, Kent Co., 2. X. 1928, C. C. Bell (DAOM, Dearness Herb.) on trunk of A. saccharum; Kanata, 18. IX. 1981, J. H. G. 6364 (DAOM 180892) on wound in trunk of living A. saccharum; London, 3. X. 1925, J. D. (DAOM) on Acer; vicinity North Bay, 22. IX. 1955, S. Gibson 55117 (DAOM 54149) on fallen Betula; Ottawa, 24. IX. 1926, I. Mounce (DAOM F675) on wounded Acer; Petawawa, 11. IX. 1939, C. G. Riley (DAOM F9565) on Acer, 24. IX. 1944, A. J. Skolko (DAOM 11917) on Acer, 17. IX. 1945, J. W. G. (DAOM 16417) on Acer; St. Lawrence Is. Natl. Park, Thwartway I., 2. X. 1975, P. Hammond (DAOM 153773) on Ulmus americana; S. Aurora, 17. X. 1931, H. S. Jackson 1149 (DAOM 80608) on Populus; Toronto, 6. XI. 1932, G. S. Bell (DAOM 50105). Quebec: Cap St. Ignace, 5. X. 1944, F. Eudore (DAOM 11955) on Acer; Chelsea, 23. IX. 1929, W. S. Odell (DAOM F8033) ; Gatineau Park, 26. IX. 1942, I. L. Conners (DAOM 11021), 27. IX. 1954, J. W. G. (DAOM 46764) on Acer; St. Aubert, 17. IX. 1946, E. Groves (DAOM 16958) on Acer, 27. IX. 1946, J. W. G. & H. A. C. Jackson (DAOM 16960) on Acer, 5. IX. 1958, J. W. G. & N. Groves (DAOM 59969) on Acer. Yukon Territory: Dezadeash, 3. VIII. 1962, W. G. Ziller (DAOM 93024 ex DAVPF 14418) on Populus trichocarpa. U. S. A.: Massachusetts :
[6] S. A. REDHEAD
Colrain, Franklin Co., 15. X. 1977, R. E. Hailing 2380 (DAOM 188111 ex FH) ring but not well documented. Additional information on the ecology and distribution on living Acer trunk. Michigan: Pellston Hills, 11. VIII. 1951, S. C. Hoare (DAOM 188003).
14. Tricholoma : The American Matsutake or Pine-mushroom (Zeller and Togashi 1934) is commonly cited as either Armillaria ponderosa (Peck) Sacc. or Tricholoma ponderosum (Peck) Singer. The rolling back of dates for valid publication at the Sydney international botanical congress now makes these names incorrect. The basionym, Agaricus ponderosus Peck (1878 : 42) was illegitimate when published, being a latter homonym of Agaricus ponderosus Persoon (1828), and therefore not available for transfer to another genus.
Agaricus ponderosus Pers. was considered to be a synonym of Agaricus columbetta Fries by Fries (1830, 1832) a distinct species presently recognized as Tricholoma columbetta (Fr.) Kummer. The epithet ponderosa did not become legitimate until Saccardo (1887) treated it in a separate genus and is properly cited as Armillaria ponderosa Sacc. (Art. 72, I. C. B. N.) with priority dating from 1887 and not 1873. In the interval, Peck (1878) apparently had become aware of Persoon's earlier homonym and proposed the new name Agaricus magnivelaris Peck as a substitute for Agaricus ponderosus Peck. Agaricus magnivelaris Peck nom. nov. is the earliest available validly published legitimate name. Murrill (1914) proposed the combination Armillaria magnivelaris (Peck) Murr. which is the correct name for the species when it is recognized as an Armillaria. However, the type of Armillaria is Armillaria mellea (Vahl. ex Fr.) Kummer as was correctly noted by Wading, Kile and Gregory (1982), and most authors do not treat Agaricus magnivelaris as congeneric with A. mellea. A more appropriate genus is Tricholoma as is accepted by this author. The following nomenclator summarizes the changes in name.
Tricholoma magnivelare (Peck) Redhead comb. nov.
basionym - Agaricus magnivelaris Peck (1878: 66) nom. nov. for Agaricus ponderosus Peck
Armillaria magnivelaris (Peck) Murrill (1914: 37)
Agaricus ponderosus Peck (1873 : 42 ; repeated 1874 : 50) nom. Meg. non Agaricus ponderosus Persoon (1828: 116)
Armillaria ponderosa Saccardo (1887: 58) ut "Peck" nom. nov. for Agaricus ponderosus Peck
Tricholoma ponderosum (Sacc.) Singer (1951 : 227) ut "(Peck) Singer"
Armillaria arenicola Murrill (1912 : 212) fide Zeller & Togashi (1934)
Tricholoma murrillianum Singer (1942 : 113) nom. nov. for A. arenicola Murr. Zeller and Togashi (1934) published the first comprehensive study of T. magnivelare noting tree associates and its occurrence in both eastern and western North America.
Kinugawa and Goto (1978) published a preliminary survey of T. magnivelare ( as Armillaria ponderosa ) in North America and indicated potentially productive areas mainly within the ranges of eastern and western hemlocks where the valuable species was suspected of occuring
Hypsizygus and Tricholoma 7
but not well documented. Additional information on the ecology and distribution has been published by Smith (1979) and Ogawa (1979). Verified collections available in the National Mycological Herbarium of Canada (DAOM) confirm areas suspected of harbouring the species by Kinugawa and Goto (1978), and in the case of the collections from Alberta and the Northwest Territories, greatly expands the known area of distribution. Mr. Gilles Patenaude noted that T. magnivelare was very abundant under mixed Pinus banksiana Lamb. and Picea glauca (Moench) Voss. It is noteworthy that no species of hemlock occurs anywhere near the northern regions (Hosie, 1973). Based on these observations it is predicted that T. magnivelare will be found across the boreal forest of Canada in suitable sandy soils.
Specimens examined :
Canada: Alberta: 22 mi. N. of Ft. McMurray, 16. IX. 1980, R. M. Danielson 3016 (DAOM 184721) under Pinus banksiana. British Columbia : Pennask Lake, 50°N 120°W, X. 1968, S. Kodama (DAOM 127345) ; North Vancouver, 5. XI. 1955, M. A. F. Waugh (DAOM 51084), 3. X. 1962, M. Pantidou (DAOM 92189) ; Wells Gray Prov. Park, Helmaker Falls, 4. IX. 1980, T. Goward 80-643 (DAOM 188295). North-west Territories, Yellowknife, 31. VIII. 1981, G. Patenaude (DAOM 187656) under Pinus banksiana and Picea on sandy burnt soil. Nova Scotia : Aldershot, 9. VIII. 1953, K. A. Harrison 2579 (DAOM 112223); Kentville, 8. X. 1931, 15. X. 1951, 9. X. 1953, K. A. H. 173, 1434, 2551 (DAOM 110623, 111829, 112197) ; Nictaux, 15. X. 1952, 17. IX. 1955, K. A. H. 1884, 5382 (DAOM 111927, 113589). Ontario: Round Lake near Pembroke, 19. IX. 1978, M. Bohart (DAOM 169162) abundant on forested sandy beach under Pinus resinosus Ait. Quebec: Lakefield, 5. XI. 1968, H. S. Cook (DAOM 116700) ; Ste. Anne de Bellevue, 18. X. 1967, H. S. C. (DAOM 124628). U. S. A.: California: Jackson State Forest near Mendocino, 29. XI. 1975, H. D. Thiers 35584 (DAOM 180686 ex SFU), also DAOM 144095, 128313, both ex SFU.
Acknowledgement — I thank Drs. J. Parmelee and J. Bissett for reviewing the manuscript.
Literature cited
Bigelow, H. E. 1976. Studies on some lignicolous Clitocybes. Mem. N. Y. Bot. Gard. 28: 9-15. Bigelow, H. E. and Barr, M. E. 1966. Contribution to the fungus flora of northeastern North America. IV. Rhodora 68: 175-191.
Bresadola, J. 1927. "Iconographia Mycologica," VI. Mediolani.
Bulliard, J. B. F. 1785. Herbier de la France. t. 5, fasc. 49-60, pls. 193-240. Paris.
Bulliard, J. B. F. 1791. Herbier de la France. t. 11, fasc. 127-132, pls. 505-528. Paris.
Bulliard, J. B. F. 1812. Histoire des champignons de la France. Deuxieme partie. Paris.
Cooke, M. C. 1882. Illustrations of British Fungi (Hymenomycetes). Vol. II. Williams and Norgate, London.
8 S. A. REDHEAD
Dennis, R. W. G. 1981. " Common British Fungi ", 2nd ed. Saiga Publ. Co., Surrey.
Fries, E. M. 1821. Systema Mycologicum. Vol. I. Lund.
Fries, E. M. 1830. Agricus synonymos in Persoonii Mycologia Europaea H et Systemate suo mycologico reconciliat. Linnaea 5: 689-731.
Fries, E. M. 1832. Index. Systema Mycologicum. Griefswald.
Hosie, R. C. 1973. Native trees of Canada. 7th ed. Can. For. Serv., Ottawa.
Imai, S. 1938. Studies on the Agaricaceae of Hokkaido. I. J. Faculty Agric., Hokkaido Imp. Univ. 43(1) : 1-178.
Imazeki, R. and Hongo, T. 1957. Coloured illustrations of Fungi of Japan. Vol. I. Hoikusha, Osaka.
Ito, S. 1959. Mycological Flora of Japan. Vol. Tr. Basidiomycetes. No. 5. Agaricales, Gasteromycetales. Yokendo Ltd., Tokyo.
Kawamura, S. 1954. Icones of Japanese Fungi. Vol. IV. Kazamashobo, Tokyo.
Kinugawa, K. and Goto, T. 1978. Preliminary survey on the "Matsutaké" ( Armillaria ponderosa ) of North America. Trans. mycol. Soc. Japan 19: 91-101.
Kuhner, R. 1938. Utilisation du carmin acétique dans la classification des Agarics leucosporés. Soc. Linn. Lyon, Bull. mem. 7: 204-211.
Lange, J. E. 1936. Flora Agaricina Danica. Vol. II. Danish Bot. Soc., Copenhagen.
Lundell, S. and Nannfeldt, J. A. 1979. Fungi Exsiccati Suecici, Praesertim Upsaliensis. Fasc. LVII-LX. Nr. 2801-3000. Reklam & Katalogtryck, Uppsala.
Miller, 0. K. 1972. Mushrooms of North America. E. P. Dutton & Co. N. Y.
Miller, 0. K. and Manning, D. L. 1976. Distribution of the lignicolous Tricholomataceae in the southern Appalachians. In: "The distributional history of the biota of the southern Appalachians. Part IV. Algae and fungi," (ed. by Parker, B. C. and Roane, M. K.) pp. 307-344. Univ. Press Virginia, Charlottesville.
Ogawa, M. 1979. Microbial ecology of 'Shiro' in Tricholoma matsutake and its allied species IX. Tricholoma ponderosa in Pseudotsuga menziesii ,Tsuga heterophylla and Pinus contorta forests. Trans. mycol. Soc. Japan 20: 370-382.
Peck, C. H. 1872. Report of the botanist. Ann. Rep. N. Y. State Mus. Nat. Hist. 24: 41-108.
Peck, C. H. 1873. Descriptions of new species of fungi. Bull. Buffalo Soc. Nat. Sci. 1: 41-72.
Peck, C. H. 1874. Report of the botanist. Ann. Rep. N. Y. State Mus. Nat. Hist. 26: 35-91.
Peck, C. H. 1878. Report of the botanist. Ann. Rep. N. Y. State Mus. Nat. Hist. 29: 29-82.
Peck, C. H. 1908. New species of fungi. J. Mycol. 14: 1-3.
Persoon, C. H. 1801. Synopsis Methodica Fungorum. Gottingen.
Persoon, C. H. 1828. Mycologia Europaea Vol. 3. Erlangae.
Pilat, A. 1935. Atlas des champignons de l'Europe. Pleurotus Fr. Ser. A. Fasc. 6-16. Praha.
Quelet, L. 1888. Flore Mycologique de la France et des pays limitrophes. Paris.
Redhead, S. A. 1979. Mycological observations : 1, on Cristulariella; 2, on Valdensinia; 3, on Neolecta. Mycologia 71: 1248-1253.
Redhead, S. A. Mycological observations, 4-12: on Kuehneromyces, Stropharia, Marasmius, Mycena, Omphalopsis, Phaeomarasmius, Naucoria, and Prunulus. Sydowia (in press).
Saccardo, P. A. 1887. Sylloge Hymenomycetum. 1. Agaricianae. Padua.
Singer, R. 1942. Type studies on agarics. Lloydia 5: 97-135.
Singer, R. 1947. New genera of fungi. III. Mycologia 39: 77-89.
ulmarius (Bull. : Fr.) Redhead(シロタモギタケ)および H. marmoreus (Peck) Bigelow(ブナシメジ)は共に優秀な食用きのこで,広葉樹の材L(倒木または生立木)にキシメジ型の比較的丈夫な肉質の子実体を形成するが,互いに子実体の外観的特徴および生態学的特徴において良く類似し,混同され易い。著者らは,1986年5 月に弘前大学で開催された日本菌学会第30 回大会において,、’シロタモギタケについて,,と題し,従来日本では,これら2種が一般に Lyophyllum ulmarium (Bull:Fr.) KUhner(和名:シロタモギタケあるいはブナシメジ)の名のもとに混同されてきたこと,確実な日本産標本に基づく両種の国内における分布および生態,並びに両種の和名の1司題について報告した。本稿はその時の内容を補足1.,てとりまとめたものである。
(ブナシメジ)多少太まり,幅は頂部で1-1.5 cm,基部で1-2.5 cm。基部はときに偽根状となる。表面は白色または基部に向ってほtまかさと同じ色を帯び,初め多少綿毛状,のち平滑。内部は充実し白色。胞子紋は淡クリーム色。
胞子(Figs. 3 and 4) は 5-6.5 x 4-5 µm,亜球形一広楕円形,やや厚膜,無色,非アミロィド,壁は鉄ィオン・酢酸カーミン処理により暗色となる。担子器は(2-)4胞子性,鉄イォン・酢酸カー ミン処理によって暗色に染まるカーミン好染性穎粒を少数もつか,またはときにこれを欠く。ひだの実質はほぼ整型。かさの表皮(Fig. 8) は平行して走る菌糸よりなり,菌糸は幅2-9 (平均4)prn,薄膜,ときに表面に帯ォリーブ色一淡黄色の沈着物をもつ。子実体を構成している菌糸には常にクランプがある。
Chapeau lisse avec marbrures innées chez tesselatus, souvent discrètes au début, peu à peu colorées, deviennent évidentes le lendemain.
Ulmarium est fibrilleux avec une micro diférente


 ブナシメジ
ブナシメジ
| Hypsizygus marmoreus (Peck) Bigelow |
ハラタケ目 シメジ科 シロタモギタケ属
写真撮影 白山 弘子

 Canada
Canada
/http%3A%2F%2Fstorage.canalblog.com%2F66%2F88%2F1309869%2F128327343_o.jpg)
/http%3A%2F%2Fstorage.canalblog.com%2F98%2F34%2F1309869%2F127348059_o.jpg)
/http%3A%2F%2Fstorage.canalblog.com%2F30%2F44%2F1309869%2F124704909_o.jpg)
/http%3A%2F%2Fstorage.canalblog.com%2F03%2F66%2F1309869%2F103591234_o.jpg)