

Macromycètes de la Nouvelle Calédonie, A. BRESINSKY
Bull. Soc. mycol. Fr., 116 (3), p. 217-229. 2000
HEBELOMA KHOGIANUM sp. nov., HYGROCYBE FIBULA sp. nov.
ET AUTRES MACROMYCÈTES DE LA NOUVELLE-CALÉDONIE
Andreas BRESINSKY *
Résumé
Description critique d'espèces de macromycètes récoltées en Nouvelle-Calédonie, dont certaines n'ont pas encore été signalées sur l'île. Deux espèces nouvelles sont proposées : Hebeloma khogianum et Hygrocybe fibula.
Abstract
Observations on fungi collected in New Caledonia are reported. Some of recorded species have previously not been observed in New Caledonia. Hebeloma khogianum and Hygrocybe fibula are established as new species.
Zusammenfassung
Es werden Beobachtungen von Pilzen aus Neukaledonien mitgeteilt. Einige der festgestellten Arten sind bisher in Neukaledonien nicht beobachtet worden. Hebeloma khogianum und Hygrocybe fibula werden als neue Arten aufgestellt.
―
|
La Nouvelle-Calédonie est un véritable trésor botanique. La richesse de sa flore en plantes vasculaires, y compris le grand nombre de paléoendémiques (parmi lesquelles la famille des Strasburgeriaceae et Parasitaxus ustus, l'unique gymnosperme parasite connue), n'est assurément pas restée sans influence sur la mycoflore locale, ce que semblent bien confirmer les études qui en ont été réalisées à ce jour. En ce qui concerne les champignons, l'île a été plus particulièrement prospectée par les mycologues français à qui revient le mérite d'y avoir identifié et décrit toute une série de genres et d'espèces du plus grand intérêt. |
* Universitât Regensburg. Fakultât fur Biologie und vorklinische Medizin. Botanisches Institut. D-93040 Regensburg (Allemagne).
218 A. BRESINSKY
Le nom générique Leratia ayant été attribué antérieurement à un genre de mousses (décrit aussi de Nouvelle-Calédonie), il a été nécessaire, eu égard aux règles de la nomenclature, de remplacer Le-Ratia par un nouveau nom, Leratiomyces (BRESINSKY & BINDER, 1998), et de procéder aux combinaisons qui s'imposaient. Parmi les travaux ayant contribué au progrès de la connaissance des macromycètes de Nouvelle-Calédonie, il convient encore de citer ceux du mycologue suisse HORAK (1979).
Une liste des taxons connus dans l'île jusqu'à une date récente a été publiée par MOUCHACCA (1998 ; ascomycètes) et HORAK & MOUCHACCA (1998 ; basidiomycètes). Dans ces deux publications, l'état de l'avancement des travaux d'herborisation mycologique est présenté de manière précise, avec la liste des déterminateurs et des récolteurs, ainsi qu'une bibliographie circonstanciée. Ces inventaires ne sont toutefois pas complets : en 1995 on pouvait, en effet, voir dans le bâtiment de l'aéroport de Nouméa une affiche en couleurs intitulée « Champignons communs de Nouvelle-Calédonie », affiche sur laquelle plusieurs des espèces illustrées, notamment Cantharellus cibarius, Hygrocybe procera et Stropharia cubensis, ne figurent pas dans la liste publiée depuis par HORAK & MOUCHACCA (1998).
Malgré toutes ces contributions, la connaissances des champignons de la Nouvelle-Calédonie est certainement encore très incomplète, comme tend à le prouver le fait que nous ayons pu y découvrir deux espèces nouvelles appartenant aux genres Hebeloma et Hygrocybe, deux genres dont, à notre connaissance, aucun représentant n'avait encore été signalé dans l'île. À l'occasion d'un bref séjour, du 13 au 26 mars 1995, nous avons eu l'occasion d'y observer, outre des représentants de ces deux derniers genres, bien d'autres espèces encore. Les polypores récoltés ont été déterminés par Leif Ryvarden (Oslo). Il s'agit des taxons suivants1 :
Amauroderma rugosum (Blume et Nees : Fr.) Torrend,
Coltricia cinnamomea (Jacq.) Murrill,
Cymatoderma elegans Jungh.,
* Datronia scutellata (Schw.) Gilb. et Ryvarden,
* Fomitopsis rhodophaea (Lév.) Imazeki,
* Ganoderma australe (Fr. : Fr.) Pat.,
Hexagonia tenuis (J. M. Book.) Fr.,
* Inonotus patouillardii (Rick) Imazeki,
* Leucophellinus hobsonii (Cooke) Ryvarden,
Microporellus obovatus (Jungh.) Ryvarden,
Microporus affinis (Blume et Nees : Fr.) Kuntze,
Microporus xanthopus (Fr. : Fr.) Kuntze,
* Polyporus gayanus Lév.,
* Polyporus philippinensis Berk.,
Pycnoporus sanguineus (L. : Fr.) Murrill,
* Rigidoporus microporus (Sw. : Fr.) Overeem,
* Rigidoporus ulmarius (Sowerby : Fr.) Imazeki,
Trametes elegans (Spreng. : Fr.) Fr.,
Trichaptum biforme (Fr.) Ryvarden.
Les récoltes appartenant à d'autres groupes se répartissaient comme suit : Auricularia delicata (Fr.) Henn., Auricularia polytricha (Mont.) Sacc., *Bertrandia astatogala R. Heim et un taxon ramené à Hygrocybe nigrescens var. brevispora (Dennis) Pegler (ces deux derniers noms sont, du reste, probablement des synonymes), *Cantharellus cibarius Fr. : Fr.,
-----------------------------------------------------------------------
1 L'astérisque (*), qui accompagne certains noms indique que ceux-ci n'apparaissent pas dans la liste de HORAK & MOUCHACCA (1998), pas même en tant que synonymes. Il s'agirait, du moins en partie, d'espèces signalées pour la première fois.
|
MACROMYCÈTES DE LA NOUVELLE-CALÉDONIE 219 * Cyptotrama asprata (Berk.) Redhead et Ginns, Pisolithus arhizus (Scop. : Pers.) Rauschert, Schizophyllum commune Fr. : Fr., *Stropharia cubensis Earle, ainsi que divers spécimens qui ont été conservés en herbier, mais dont la détermination n'a pas été possible; il s'agit de représentants de genres tels que *Galerina et *Pluteus, genres dont aucun taxon n'est répertorié dans l'article de HORAK & MOUCHACCA (1998). Les spécimens secs de toutes les espèces citées, ainsi que le matériel correspondant aux taxons faisant l'objet du présent article, ont été déposés dans l'herbier de la Botanische Staatssammlung München (M). Certaines récoltes concernent des espèces qui, à ce jour, n'ont jamais été décrites; ce sont elles qui seront discutées et nommées dans ce qui suit. Hebeloma khogianum, sp. nov. Pileus 8 cm, pileocutis primo modice lubrica, leviter nitida, propius marginem magnis accumbentibus, in ipso margine praeacutis squamis fibrillosis dependentibus instructa. Color pileocutis ochraceus, propius marginem flavescentes. Lamellae sinuatae decurrentes, colore carneo vel ex rufo ochraceo, non argillaceo. Acies lamellarum et margine albido et maculis hic illic obscuris instructae. Stipes 10 x 1,3 cm, basi breviter radicata, quae basis supra radicem non insigniter est clavata, summa in parte anulo instructus, coloreflavo vel argillaceo, infra anulum fibrillis et squamis accumbentibus tectus. Caro primo odore fugaci dulci et martipanis simillimo, deinde odore fungorum consueto aut odoris expers. Nascitur in humo prope semitam quandam in monte Khogis insulae Novae Caledoniae. Sporae latae, ellipsoideae vel amygdaliformes, verrucosae simulque crasse tunicatae, sub microscopio (colon pulveris sporarum non exquirebatur) ex brunneo ochraceae, 7-8 (-9,5) x5-6 (-7) pm. Lectum in monte Khogis, die 23. Maii 1995; holotypus in herbario Monacensi (M) servatur. Chapeau : 8 cm, campanulé à campanulé-obtus, puis convexe, à marge et marginelle involutées, d'abord légèrement lubrifié, mais vite sec, un peu luisant, revêtu d'écailles fibrilleuses brun ochracé à brun argilacé, pointues et appendiculées au bord, larges et apprimées à l'ambitus où la couleur jaune paille du fond domine, et s'amenuisant en fines squamules vers le centre qui, du fait de la densité de cette formation, apparaît brun ochracé. Lames : émarginées-uncinées, assez larges (6 mm), avec plusieurs longueurs de lamellules, serrées, contiguës au fond des sinus, ocre rougeâtre ou d'un incarnat particulier (non brun argilacé !) ; arête sinueuse, ébréchée, blanche par une pruine floconneuse, tachée de brun sombre par endroits. Stipe : 10 x 1,3 cm, cylindracé, à base atténuée et courtement radicante sous un vague renflement claviforme (atteignant x 1 cm), paille clair, plus ou moins flocculeux et irrégulier-ridé au sommet, pourvu, en haut, d'un anneau brun argilacé clair, fibrillo-membraneux, peu adhérent (sur les exemplaires suffisamment frais), qui, plus bas, passe à des fibrilles ou squamules apprimées de même couleur. |
220 A. BRESINSKY
Chair blanche, particulièrement dans le chapeau, d'un ochracé sale dans le stipe sous la surface et uniformément dans la partie radicante, molle dans le chapeau où elle peut atteindre 1 cm d'épaisseur, ferme dans le stipe. Odeur fugace et suave de pâte d'amandes, puis insignifiante ou banale. Saveur amarescente à presque douce.

Caractères microscopiques : spores' 7,0-8,0 (-9,5) x 5,0-6,0 (-7,0) µm, soit en moyenne 7,07 x 5,51
largement ellipsoïdes-amygdaliformes, à face interne aplatie et à face externe fortement bombée, renfermant
une ou plusieurs guttules, ocre brunâtre pâle sous le microscope (la couleur de la sporée n'a pas pu être déterminée), verruqueuses, à paroi épaisse et montrant un myxosporium hyalin. Arête des lames stérile, densément couverte de cheilocystides cylindriques à légèrement claviformes, 40 x 7 x 3 µm, exsudant, dans une solution aqueuse de potasse, des gouttelettes huileuses.
Habitat et lieu de récolte : à terre, en bordure d'un chemin, non loin d'un groupe de Gleichenia, au voisinage de Myrtaceae, non associé aux cavernes ou repaires d'animaux terrestres. Forêt tropicale humide du mont Khogis (Nouvelle-Calédonie), le 23 mai 1995 ; leg. A. Bresinsky.
-----------------------------------------------------
2 Les mensurations extrêmes citées entre parenthèses proviennent d'une unique spore, vraisemblablement anormale.
MACROMYCÈTES DE LA NOUVELLE-CALÉDONIE 221

222 A. BRESINSKY
DISCUSSION
La simple vue du carpophore, jointe à l'odeur de pâte d'amandes, pour fugace qu'elle soit, suffit à évoquer l' Hebeloma radicosum de l'hémisphère boréal, et, le fait est, qu'il est permis de voir dans notre hébélome néocalédonien un représentant du sous-genre Myxocybe caractérisé par des relations de parenté étroites avec H. radicosum.
Une autre espèce décrite de l'hémisphère austral, Hebeloma victoriense Holland et Pegler, présentait déjà un cas semblable (HOLLAND & PEGLER, 1983). En raison de la couleur de la sporée, Holland et Pegler étaient cependant enclins à placer leur taxon plutôt dans le sous-genre Porphyrospora. Mais si, comme nous le pensons, on doit accorder une importance plus grande à la morphologie (stipe radicant, anneau) qu'à la couleur de la sporée, il est alors préférable de classer H. victoriense, ainsi que H. khogianum (dont la sporée, rappelons-le, n'a pu être obtenue), dans le sous-genre Myxocybe. Chez H. victoriense, la forte ressemblance du carpophore et des spores avec ceux d'H. radicosum mène à envisager une parenté très
proche, si ce n'est même la conspécificité de ces deux taxons.
Hebeloma victoriense, qui, jusqu'à présent, n'est connu que du sud de l'Australie, se distingue de H. khogianum par la taille et la forme de ses spores. En effet, la longueur minimale de celles du premier correspond dans l'ensemble à la longueur maximale de celles du second, alors que la largeur maximale est la même dans les deux cas (les spores de H. victoriense ont 8-10 x 4-6 µm, celles de H. khogianum 7-8 x 5-6 µm). On note également une différence dans la longueur et la largeur moyennes des spores (8,9 x 5,15 µm chez H. victoriense, contre 7,07 x 5,51 µm chez H. khogianum). Ceci reflète bien les
nettes différences observées dans la forme des spores qui, chez H. khogianum, ne sont plus amygdaliformes, mais largement ellipsoïdes. Par ailleurs, H. khogianum diffère de H. radicosum de manière très comparable.
Selon des données trouvées dans la littérature, les spores de H. radicosum auraient 8-10 x 5,5-6 µm. Nos propres mesures, effectuées sur du matériel récolté en Bavière (dans la région de Ratisbonne), ont fourni 9-10 x 5-6 µm (moyenne : 9,55 x 5,57 µm). Comme dans le cas de H. khogianum et de H. victoriense, les spores d'H. radicosum montrent un myxosporium hyalin.
Chez H. radicosum, comme chez H. khogianum, des fragments de lamelle, ainsi que les spores, observés au microscope, après montage dans une solution aqueuse de potasse, exsudent dans ce milieu des gouttelettes huileuses. Les verrues constituant l'ornementation sporale semblent un peu plus marquées chez H. radicosum que chez H. khogianum. La comparaison de figures représentant des spores de ces deux derniers (fig. 1) permet de mieux se rendre compte de ce qui les différencie. De ces constatations résulte, en accord avec l'illustration fournie par HOLLAND & PEGLER (1983), que H. khogianum diffère de H. victoriense et de H. radicosum de manière très semblable dans les deux cas. La mesure du contenu en A.D.N. du noyau sporal au moyen du procédé DAPI (WITTMANN-BRESINSKY & BRESINSKY, 1989), fournit une teneur relative de 8,96 + 2,7 pour H. khogianum, contre 24,79 +
223
4,52 chez l'H. radicosum européen3. Le noyau sporal d'H. radicosum a donc un contenu en A.D.N. sensiblement trois fois supérieur à celui de H. khogianum, ce qui peut encore être interprété en disant que le génome du premier est significativement plus gros (trois fois) que celui du second. Il se peut donc que chez H. khogianum nous soyons en présence du « taxon originel » monoploïde et chez H. radicosum du dérivé triploïde correspondant immigré dans l'hémisphère boréal.
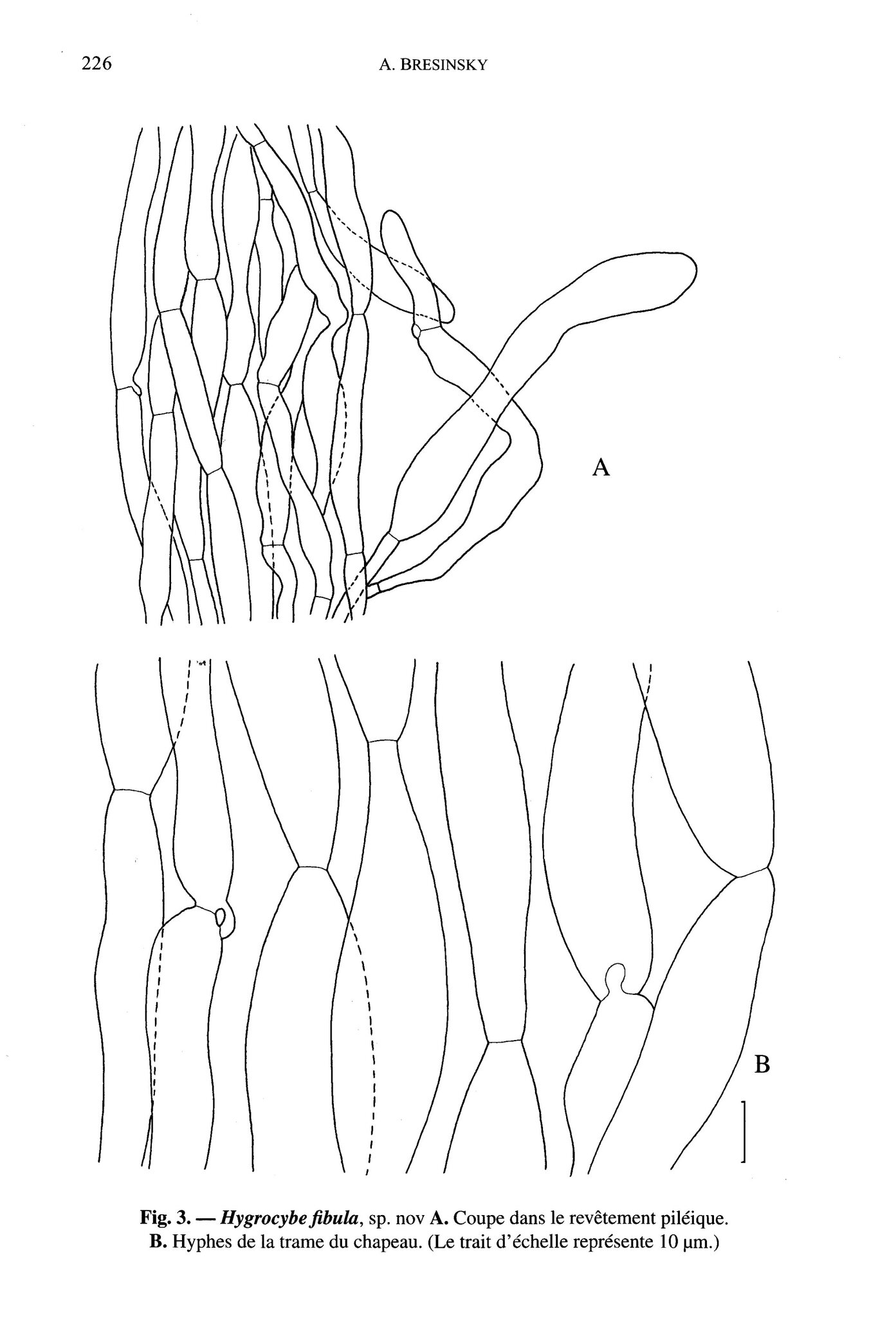
Hygrocybe fibula, sp. nov.
Pileus ad 2 cm latus, molliter convexus, centro applanato aut leviter depresso, margine crenulato et undulato, colore obscure cerasino, superficie sicca, languida, furfuracea vel squamosa, ipsis quoque squamulis obscure rubidis. Lamellae alte decurrentes, distantes, albido-ochraceae, de parte intima rufescentes ; acies lamellarum subcrenulata ac inaequalis. Stipes 5-8 x 0,2 cm, prope basim ad similitudinem fibulae attenuatus, cerasinus excepta basi paulum lucidiore et aurantiaca. Caro ubique cerasina, sapore miti, sine odore.
Sporae 8,5-9,0 x 60-7,0 pm, non strangulatae, majore ex parte uninucleatae. Basidia 40-50 x 8-10 ,um, aut 2
aut 4 sterigmatis apparata. Trama lamellarum de elementis constat curtis, latius ellipsoideis vel paene sphaeroideis.
Lecta ad Flumen Caeruleum (Rivière-Bleue), die 26. Maii 1995; holotypus in Herbario Monacensi (M) servatur.
Le genre Meiorganum, proche des bolétales, fut publié par R. Heim en même temps que le rare Meiorganum neocadelonicum, à partir de récoltes faites sous les Nothofagus des magnifiques forêts tropicales de Rivière-Bleue. Une telle découverte vient s'intéger dans le cadre de celle, également récente, d'une série de phanérogames néocalédonniennes. Cette remarque vaut en particulier pour le genre Nothofagus, signalé relativement tard en Nouvelle-Calédonie et où il s'avéra exclusivement représenté par de nouvelles espèces, jusqu'alors complètement inconnues. C'est dans cette même région de Rivière-Bleue, qu'il nous a été donné d'observer, se développant en compagnie de Cantharellus cibarius et Pisolithus arhizus, un petit Hygrocybe rouge, manifestement non encore signalé et qui donc restait à décrire comme nouveau.
Chapeau : 2 cm, d'abord campanulé-obtus, déployé-convexe, puis plan-convexe et plus ou moins aplati ou même légèrement déprimé au disque, crénelé-onduleux à la marge, sec, mat, rouge cerise intense (10 — 11 B 8 du code de couleurs de KORNERUP & WANSCHER, 1975) et revêtu de squamules ou excoriations concolores.
Lames : fortement décurrentes, commençant leur descente près de la marge piléique, entremêlées de courtes et étroites lamellules, distantes, ménageant un écart au fond des sinus, à arête crénelée ou irrégulière, ocre pâle à ochracé sale, y compris en vue de profil, mais avec un reflet rougeâtre au fond, en vue frontale.
Stipe : 5-8 x 0,2 cm, cylindracé, plein, élastique et tenace, atténué vers la base, en forme de fibule, rouge cerise, concolore au chapeau, sauf à la base, sec, mat, subtilement strié sous la loupe, mais glabre et lisse à l'oeil nu, manifestement non pruineux.
3 Cette teneur est exprimée ici en pourcentage de la teneur en A.D.N. d'un étalon.
224 A. BRESINSKY
PLANCHE II

Chair : entièrement rouge cerise intense, pâlissant très lentement des tons ochracé rougeâtre
au coeur du stipe et du chapeau. Odeur et saveur banales ou insignifiantes.
Caractères microscopiques : spores 8,5-9 (-11) x 6-7 (-8) µm, largement ellipsoïdes, uninuclées4; basides à 2 ou 4 stérigmates, 40-50 x 8-10 µm; trame des lames constituée d'hyphes brièvement articulées, aux articles boursouflés, larges (x 2-35 µm), ellipsoïdes à subglobuleux, en mélange avec des hyphes grêles aux articles étirés ; revêtement piléique constitué d'hyphes couchées parallèlement, avec, çà et là, une boucle de petite dimension aux cloisons; squamules constituées d'hyphes ascendantes, organisées en une sorte de trichoderme et à articles terminaux (x 13-18µm) claviformes ou ampullacés.
Photographie : A. Bresinsky
Hygrocybe fibula Bresinsky, sp. nov.
Lieu de récolte : dans le parc forestier de Rivière-Bleue (Nouvelle-Calédonie), le 26 mai 1995 ; leg. A. Bresinsky.

DISCUSSION
Cette espèce, décrite ici comme nouvelle, présente, du fait de ses lames fortement décurrentes, une lointaine ressemblance avec H. cantharellus (Schw.) Murrill et H. firma var. trinitensis Dennis. La variété typique, H. firma Corner var. firma, diffère de notre champignon par sa dimorphie sporale 5. Cette particularité du type de H. firma,
en soi très caractéristique, ne se retrouverait pas dans la var. trinitensis, tout au moins si l'on se limite à ce qu'en
dit Dennis dans la diagnose originale, d'où la nécessité de discuter séparément des différences entre H. fibula et
H. firma var. trinitensis.
4 Nous n'avons pu détecter aucune spore binuclée.
5 Une telle dimorphie se présente lorsque, eu égard aux dimensions, les basides, et donc aussi les spores, se répartissent suivant deux classes distinctes (CORNER, 1936).
L'un des carctères différentiels utilisés par DENNIS (1953, 1970) dans ses clés pour séparer H. cantharellus de
H. firma var. trinitensis, à savoir la couleur du stipe, n'est pas d'une grande aide ici, car celui de H. cantharellus, contrairement à ce qu'indique Dennis par erreur, est également rouge. Les trois taxons impliqués (H. cantharellus, H. firma var. trinitensis et H. fibula) doivent donc être examinés plus en détail afin de préciser ce qui les différencie vraiment les uns des autres. Les spores de H. firma var. trinitensis (11-13 x 5-6 µm) sont plus longues et plus
étroites que celles de H. fibula (8,5-9 x 6-7 µm). Selon des travaux plus récents de PEGLER & FIARD (1978), H. firma var. trinitensis doit maintenant être considéré comme une espèce autonome, H. trinitensis, distincte de H. firma. Dans le même temps, la diagnose de H. trinitensis doit être amendée de manière à y faire figurer une dimorphie sporale que ce dernier partage donc avec H. firma. Les dimensions sporales à prendre en considération pour H. trinitensis sont maintenant 10-13 x 6- 7,51.1,1µm pour le groupe correspondant au plus grand mode et 7-9 x 4,5-5,5 µm pour l'autre groupe. Les spores d'H. fibula (8,5-9 x 6-7 µm) ne s'accordent donc avec aucun de ces deux groupes, pour ce qui est des dimensions. Chez H. procera (Stevenson) Horak, donné comme espèce commune en Nouvelle-Calédonie sur l'affiche dont il a été question plus haut, les spores mesurent 11-17 x 7-10
µm (selon HORAK, 1973) et, là encore, diffèrent nettement tant en ce qui concerne la forme que les dimensions.µm
Les spores de H. cantharellus (6-9 x 4-6 µm), du moins quant à leurs longueurs, se rapprochent davantage de celles de H. fibula, mais elles continuent néanmoins à en différer par la largeur. De plus, H cantharellus possède des spores binucléées, tandis que celles de H. fibula sont uninucléées. Parmi les hygrocybes comparables, H miniata (Fr.) P. Kumm., et l'espèce voisine H. calciphila Arnolds, ont des spores également uninucléées. Hygrocybe fibula, dont les spores, par leur forme et, plus encore, par leur dimension, se rapprochent de celles de H calciphila, diffère de ce dernier par ses carpophores plus petits, aux lames pâles et plus nettement décurrentes. Mais il faut souligner encore une fois à cette occasion que H. fibula et miniata (ou calciphila) ont un habitus tout à fait différent. H. helobia diffère non seulement par l'habitus, la coloration des spores et leurs dimensions, mais encore par le caractère binucléé de ces dernières. La variété typique de H. firma, c'est-à-dire H. firma var. firma, montre également des spores binuclées, et ce, dans les deux groupes résultant du dimorphisme.
De même, en raison de la teneur en A.D.N. de son noyau sporal, une position particulière semble devoir être réservée à H. fibula. Se situant aux environs de 25 %, le contenu en A.D.N. relatif de sa spore est inférieur à celui de H cantharellus (45-88 %), de H. firma (72-131 %), de H. miniata (47-88 %) et de H. helobia (41-59 %). Même si, à cause de la forte fluorescence d'arrière plan, l'écart-type chez Hygrocybe fibula reste relativement élevé en avoisinant 25 %, la différence n'en reste pas moins significative. Ajoutons, pour terminer, que les caractéristiques de la trame des lames, constituée d'articles très larges ou globuleux, parlent également en faveur de la séparation spécifique de H. fibula entreprise ici.
228 A. BRESINSKY
REMERCIEMENTS
Je tiens à remercier Fatima Thrun (Munich) pour la traduction en français de l'article et O. Raith (Ratisbonne) pour la traduction en latin des diagnoses des espèces nouvelles qui y sont publiées. Ces remerciements vont aussi à I. Schônberger (Ratisbonne) pour les mesures de la teneur relative en A.D.N. du noyau sporal d' Hebeloma khogianum et à H. Besl (Ratisbonne) pour son aide bibliographique. Il m'est agréable de remercier tout particulièrement Leif Ryvarden (Oslo) pour la détermination des aphyllophorales citées dans ce travail, ainsi que M. Beisenherz (Ratisbonne) pour l'aide qu'il m'a apportée lors de l'exécution des différentes analyses microscopiques et pour son assistance technique lors des mesures de la teneur en A.D.N. des noyaux d' Hygrocybe fibula.
BIBLIOGRAPHIE
BINDER, M., H. BESL, & A. BRESINSKY. 1997. — Agaricales oder Boletales ? Molekularbiologische Befunde zur Zuordnung einiger umstrittener Taxa. Z. Mykol., 63, p. 189-196.
BRESINSKY, A., & M. BINDER. 1998. - Leratiomyces nom. nov. ftir eine bislang nicht gültig beschriebene Gattung der Strophariaceae (Agaricales) aus Neukaledonien. Z. Mykol., 64, p. 79-82.
CORNER, E. J. H. 1936. — Hygrophorus with dimorphous basidiospores. Trans. Brit. Mycol. Soc., 20, p.-157-184.
DENNIS, R. W. G. 1953. — Some West Indian collections referred to Hygrophorus Fr. Kew bull., Addit., ser., 3, p. 253-268
DENNIS, R. W. G. 1970. — Fungusflora of Venezuela and adjacent countries. Cramer, Lehre. HEIM, R. 1951. — Notes sur la flore mycologique des terres du Pacifique Sud. Le genre néo-calédonien Le-Ratia. Rev. mycol. [Paris], 16, p. 154-158.
HEIM, R. 1965. — Les Meiorganés, phylum reliant les Bolets aux Polypores. Rev. Mycol. [Paris], 30, p. 307-329.
HEIM, R. 1967. — Notes sur la flore mycologique des terres du Pacifique Sud. Rev. mycol. [Paris], 32, p. 3-15.
HEIM, R. 1968. — Notes sur la flore mycologique des terres du Pacifique Sud. Rev. mycol. [Paris], 33, p. 137-152.
HEIM, R. 1971. — The interrelationships between the Agaricales and Gastromycetes. DansEvoluti on in the higher Basidiomycetes, publié sous la direction de R. H. Petersen. Knoxville.
HOLLAND, A. A., & D. N. PEGLER. 1983. - Hebeloma victoriense and the genus Metraria. Trans. Brit. Mycol. Soc., 80, p. 157-186.
HORAK, E. 1973. — Fungi agaricini novazelandiae. I-V. Beih. Nova Hedwigia, 43, p. 1-200.
HORAK, E. 1979. — Paxilloid Agaricales in Australasia. Sydowia, 32, p. 154-166.
HORAK, E., & J. MOUCHACCA. 1998. - Annotated checklist of New Caledonian Basidiomycota. I. Holobasidiomycetes. Mycotaxon, 68, p. 75-129.
KORNERUP, A., & K. A. WANSCHER. 1975. — Methuen, Taschenlexikon der Farben. 2. Aufl. Zürich, Giittingen.
MOUCHACCA, J. 1998. — Ascomycetes described from New Caledonia, South Pacific region. Mycotaxon, 67, p. 99-121.
MACROMYCÈTES DE LA NOUVELLE-CALÉDONIE 229
PATOUILLARD, M. N. 1907. — Le Ratia, nouveau genre de la série des Cauloglossum. Bull. Soc. mycol. France, 23, p. 50-52.
PATOUILLARD, M. N. 1909. — Champignons de la Nouvelle-Calédonie (suite). Bull. Soc. mycol. France, 25, p. 129-134.
PEGLER, D. N., & J. P. FIARD, 1978. - Hygrocybe sect. Firmae in Tropical America. Kew bull., 32, p. 297-312.
WITTMANN-BRESINSKY, B., & A. BRESINSKY, 1989. - Quantitative differences of nuclear DNA contents in Paxillaceae compared with other families of Boletales. Mycol. Res., 93, p. 282-291.
/http%3A%2F%2Fstorage.canalblog.com%2F98%2F79%2F1309869%2F119938886_o.jpg)
/http%3A%2F%2Fstorage.canalblog.com%2F47%2F79%2F1309869%2F122644164_o.jpg)
/http%3A%2F%2Fstorage.canalblog.com%2F01%2F34%2F1309869%2F120907545_o.jpg)
/http%3A%2F%2Fstorage.canalblog.com%2F45%2F05%2F1309869%2F105471448_o.jpg)